7. Рост, размножение и экологическая стратегия
С точки зрения экологии подводные оазисы представляют собой совершенно оригинальное явление, не имеющее эквивалента на нашей планете:
- Гидротермальная среда имеет столь специфические физико-химические характеристики, что те из обитающих в ней видов, которые действительно к ней приспособились и количественно составляют основную часть сообщества, не могут существовать ни в каких других условиях.
- Гидротермальная среда принципиально нестабильна: в масштабе отдельного излияния продолжительность жизни гидротермального источника исчисляется десятками лет, самое большее столетием; в региональном масштабе (от нескольких десятков до нескольких сотен километров вдоль хребта) продолжительность жизни гораздо больше, но все же ограниченна.
Каким же образом гидротермальное население приспособилось к этой среде? Другими словами, какова та адаптивная стратегия, которая позволяет этим сообществам выживать на уровне особи (то есть каковы адаптации, имеющие отношение к поведению и физиологии) и на уровне популяции (выживание популяции, если не говорить об индивидуальных возможностях, зависит от того, как происходит воспроизводство особей рассматриваемой популяции)?
В экологической теории, построенной на наземных моделях, но полностью приложимой к морской среде, различаются две противоположные адаптивные стратегии:
- Стратегия, приспособленная к жизни в нестабильных средах, где физико-химические условия меняются в широких пределах; соответствующие виды очень толерантны по отношению к физико-химическим условиям, имеют высокую скорость роста, раннее половое созревание и большую способность к расселению. То есть, в антропоморфных терминах, эта стратегия заключается в том, чтобы как можно быстрее извлекать пользу из возникших в определенной среде на короткое время благоприятных условий.
- Стратегия, приспособленная к жизни в стабильных средах, где физико-химические условия постоянны или меняются незначительно с течением времени; соответствующие виды узко приспособлены к особенностям своей среды, имеют малую скорость роста, позднее половое созревание и слабую способность к расселению.
В случае гидротермальных сообществ можно было ожидать преобладание видов первой группы, которых экологи называют r-стратегистами, в отличие от видов второй группы - К-стратегистов.
Хотя современные знания еще очень малы, можно отчасти ответить на поставленный вопрос в отношении крупных видов, лучше всего приспособившихся к гидротермальной среде.
Биологи довольно давно занялись исследованием скорости индивидуального роста двух крупных двустворчатых моллюсков, пытаясь выяснить, соответствует ли их высокая биомасса столь же высокой продукции. С большим или меньшим успехом были использованы различные методы. Для батимодиолы с Галапагосского рифта в 1979 году удалось использовать прямой метод, который состоит в мечении особей путем нанесения меток на раковины с помощью зубоврачебной фрезы, управляемой манипулятором подводного аппарата. Через известный промежуток времени меченых животных достают (так называемый метод пометки - поимки). Помеченные батимодиолы были выловлены через 294 дня. Скорость роста моллюска длиной 18 см в среднем равна 1 см в год; она высока у молодых особей и убывает с возрастом. У калиптоген первоначальный расчет по скорости распада естественных радиоизотопов (серии 230U и 232Th) элементов, входящих в состав известковой раковины - измеряли соотношения радиоизотопов в разных точках по оси максимального роста раковины, - дал следующий результат: скорость роста равна 5 см/год для первых 6 см длины раковины и 2-3 см/год на каждые последующие 2 см, средний прирост за всю жизнь 4 см/год, что является удивительно высоким результатом. Другие методы, основанные на изучении микроструктуры раковины и использовании отношений изотопов 18O /16O и 13С / 12С в карбонате кальция раковин калиптоген, дали более низкие значения скорости роста и, соответственно, более значительный возраст. В то же время эти методы позволили выявить определенную ритмичность роста, которая, вероятно, зависит от изменений активности гидротермальных излияний. Самые последние подсчеты вводят поправку на скорость растворения раковин: действительно, на такой глубине кислотность разбавленного гидротермального флюида достаточна, чтобы за несколько десятков лет полностью растворить раковины двустворок, и растворение начинается еще при жизни моллюска. При изучении калиптогены этими методами результаты оказались ближе к тем, что были получены на батимодиолах, чем очень высокие значения, которые дал метод измерения распада радиоизотопов, входящих в состав раковины: 1,2 см/год для особей длиной 12,5 см, то есть моллюсков среднего размера. Даже если между американскими и французскими специалистами имеются некоторые разногласия, можно ориентироваться на следующие средние (значения скорости роста: 1 см/год для батимодиолы и 0,8-1,1 см/год для калиптогены, причем максимальный возраст у калиптогены выше, чем у батимодиолы, и составляет 20-30 лет.
Таким образом, гидротермальные двустворчатые моллюски растут в два-три раза быстрее, чем живущие на сравнимых глубинах другие виды двустворок, размер которых не превосходит 1-1,5 см. В случае батимодиолы наблюдаемая скорость роста сравнима со скоростью роста прибрежных мидий. Другими словами, когда количество пищи не ограничено, митилиды растут со скоростью, которая не зависит от особенностей гидротермальных источников.
Достоверных данных, позволяющих оценить скорость роста каких-либо беспозвоночных с мягким телом, помимо двустворок, сейчас практически нет. Некоторые наблюдения, сделанные на 13° с. ш. с интервалом в два года, похоже, свидетельствуют о высокой скорости роста трубок рифтий (но, разумеется, рост самих животных остается неизвестным, поэтому наблюдения следует повторить и дополнить).
Вторая важная адаптивная особенность касается Демографической стратегии: размножения видов и существования расселительных стадий. Имеющиеся немногие результаты касаются, во-первых, батимодиол из излияний на Галапагосском рифте и на 13° с. ш. и, во-вторых, некоторых видов, для которых есть отрывочные данные. В целом результаты несколько противоречивы.
Яйца батимодиолы довольно мелкие (60 мкм в диаметре), и она выметывает их по одному (у прибрежных мидий яйца выметываются пакетами по нескольку десятков). У батимодиол есть планктонная личинка, обеспечивающая распространение вида. Это, правда, требует комментария. Дело в том, что пока никто не видел личинок батимодиол. Но изучение макушек раковин (то есть, самой молодой части раковины) взрослых особей в сканирующем электронном микроскопе позволяет установить существование двух последовательных личиночных стадий, хорошо известных специалистам под названием продиссоконхи I и II. Продиссоконха I имеет в длину 95 мкм, продиссоконха II - более 400 мкм. Специалисты по двустворчатым моллюскам считают, что маленькие продиссоконхи I соответствуют пелагическим личинкам, которые долгое время живут и питаются в планктоне (планктотрофные личинки), а большие продиссоконхи I (130-500 мкм) соответствуют личинкам с укороченной (или отсутствующей) пелагической стадией (лецитотрофные личинки). Маленьким продиссоконхам I соответствуют гладкие (без скульптуры) и крупные продиссоконхи II, и наоборот. Следовательно, у батимодиолы есть пелагическая планктотрофная личинка, способная жить в планктоне много недель, а может быть даже несколько месяцев, при температуре 2 °С. Тогда при скорости течения 10-20 см/с возможно расселение таких личинок на расстояния во многие тысячи километров; эта особенность свойственна видам с r-стратегией, что, однако, противоречит малому количеству откладываемых яиц. Но ведь личинку батимодиолы еще никто не видел!
Примененный к калиптогене, тот же метод дал противоположный результат: продиссоконха II полностью отсутствует, что свидетельствует об очень короткой или вовсе отсутствующей пелагической стадии и, вследствие этого, крайне низких расселительных способностях. Точно так же нет пелагической личиночной стадии у большинства колпачковидных брюхоногих моллюсков (у брюхоногих говорят о протоконхе I и II, но соотношения те же самые: протоконха I - это первичная раковина, формирующаяся во время эмбрионального развития, протоконха II - раковина, строящаяся во время пелагической жизни). У недавно описанного маленького гребешка, ставшего одним из примеров живого ископаемого (см. гл. 8), наоборот, есть продиссоконха I маленького размера и планктотрофная личинка.
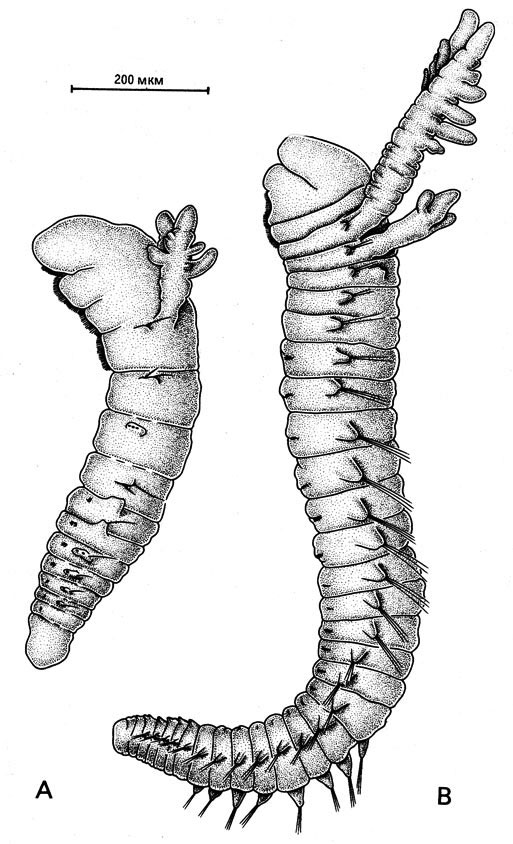
Рис. 29. Очень молодые особи Paralvinella grasslei, которые перемещаются, ползая по субстрату. Способ распространения альвинеллид и, в частности, помпейских червей с одного местонахождения на другое пока совершенно не ясен
Яйца рифтии имеют размер 70-100 мкм, а самые маленькие из собранных особи, уже имеющие облик взрослых, не превышают 2 мм. Можно предполагать, исходя из того что мы знаем о развитии погонофор, что между этими двумя стадиями существует личинка, покрытая редкими ресничками, которая ползает по субстрату и имеет весьма ограниченные способности к распространению. Точно так же самая молодая стадия, известная для помпейских червей, имеет длину менее 1 мм и насчитывает примерно 12 сегментов. По аналогии с близкими видами можно полагать, что у этих животных нет расселительной личиночной стадии. Однако геологи наблюдали роение червей, похожих на помпейских, которые плавали в воде в нескольких метрах надо дном, и по крайней мере одну особь Alvinella caudata, плававшую в воде, наблюдал и точно определил биолог. Может быть, это и есть механизм расселения вида на взрослых стадиях, хорошо, кстати, известный экологам для других видов? Наконец, в жаберной полости Bythograea нашли взрослых червей другого вида, Amphisamytha galapagensis: можно полагать, что таким образом краб переносит их из одного местонахождения в другое.
Эти несколько наблюдений зоолога не позволяют установить связи между разными гидротермальными излияниями одного района, идет ли речь о возрасте населения сообщества (то есть о дате начала заселения) или о происхождении первых особей каждого вида, которые постепенно заселяли излияние. Методы экологической генетики, которые, в частности, позволяют установить существование одной или двух генетически различных популяций в одном сборе, могут оказаться чрезвычайно полезными для развития этих направлений. Пока лишь батимодиолы из двух местонахождений на Галапагосском рифте стали объектами генетического исследования американских ученых, еще не опубликованного.
Вопрос о распространении и выживании гидротермальных сообществ реально стоит в малых масштабах времени и пространства, порядка сотни лет и десятка километров. Проблема биогеографического распространения гидротермальных сообществ и изменений видового состава, которые отмечаются при сравнении двух крупных совокупностей излияний, расположенных на севере (хребты Хуан-де-Фука и Эксплорер) и на юге (21 и 13° с. ш. на Восточно-Тихоокеанском поднятии и Галапагосском рифте) от Орегонской зоны субдукции, ставит другие вопросы, которые касаются уже эволюции. Разрыв между этими двумя большими совокупностями произошел примерно 25 млн. лет назад. Канадская исследовательница Танниклиф считает, что степень дифференциации гидротермальной фауны этих двух областей недостаточно велика для столь большого промежутка времени. Следует ли из этого, что гидротермальные излияния - очень консервативная среда, скорость эволюции в которой мала, или же что существует еще не известная географическая непрерывность распространения, а может быть, просто наши зоологические познания еще слишком несовершенны? И здесь тоже метод эволюционной генетики, позволяющий восстановить степень родства между близкими видами, поможет лучше понять механизм преобразования видов и механизмы организации разных типов сообществ.
Самый предварительный вывод, возможно, состоит в том, что если в локальном масштабе пространственно-временная изменчивость гидротермальной среды очень велика, то в масштабе региональном, и даже мировом, и в ходе геологического времени гидротермальная среда очень постоянна. Эта двойственность, вероятно, объясняет, почему адаптивные стратегии сложнее, чем это может показаться априори, исходя из чисто локальных оценок.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'