Рациональная эксплуатация хозяйственно-ценных популяции
Очень важный метод повышения продуктивности популяций промысловых организмов - это их рациональная эксплуатация. Как говорилось выше, популяции любого вида организмов, как растительных, так и животных - это непрерывно работающий трансформатор энергии. Это система, получающая энергию из внешнего мира и перерабатывающая ее согласно своей видовой специфике, строящая из нее свою биомассу и отдающая энергию обратно во внешний мир. Естественно, что у разных видов организмов и при разных условиях интенсивность этого процесса неодинаковая и характеризуется у каждого вида своей качественной спецификой. При разных режимах жизни популяции соотношение энергии, идущей на построение ее биомассы, т. е. тела слагающих ее организмов, и на поддержание жизни весьма различное.
Задача в том и заключается, чтобы разработать такие режимы "работы" популяции хозяйственно-ценных организмов, которые позволяли бы при минимуме потребляемых популяцией ресурсов получать максимальное количество хозяйственно-ценной продукции нужного ассортимента и качества.
В общем виде задача заключается в том, чтобы при имеющихся в водоеме кормовых ресурсах получать максимум рыбной продукции наиболее высокого качества. Для этого необходимо в первую очередь, чтобы промысловые организмы использовали максимум кормовых ресурсов, причем наиболее эффективно, т. е. при минимальном кормовом коэффициенте. Чем большее количество рыбы можно получить в виде второго звена трофических цепей, т. е. растительноядных рыб, тем обычно выше общая продуктивность водоема (см. выше).
Однако добиться того, чтобы вся съедаемая в водоеме растительность непосредственно шла на построение тела хозяйственно-ценных организмов, практически не удается. Если бы это можно было сделать, то, конечно, прирост продукции весьма возрос бы. Как видно из табл. 104 (Никольский, 1956), в разных бассейнах нашей страны улов можно было бы весьма увеличить, если бы получать всю рыбную продукцию в виде второго звена пищевых цепей.

Таблица 104. Возможная рыбная продуктивность при получении хозяйственно-ценной продукции в виде растительноядных (второе звено пищевых цепей) и бентосоядных (третье звено) рыб по сравнению с современной продуктивностью, принятой за 100%
Однако при этом надо учитывать, что если в континентальных водоемах за счет растительноядных рыб можно получить весьма высокоценную продукцию, то в океане особо ценных видов рыб-фитофагов немного, а основную массу их составляют относительно мелкие виды. В океане наиболее ценными являются животноядные бентофаги или хищники.
Естественно, что в северных широтах относительная роль растительности, потребляемой хозяйственно-ценными организмами, будет много меньше, чем в водоемах низких широт. При построении рационального рыбного хозяйства на том или ином водоеме очень важно правильно спланировать видовой состав уловов. Это, конечно, не только биологическая, но в значительной мере и экономическая проблема. Экономист должен решить, что в настоящее время важнее: получить, например, с Азовского моря 1 кг судака или 5-7 кг бычков и тюльки; либо с озер Белоруссии 1 кг щуки или 7-10 кг мелкой плотвы и окуня. В зависимости от того, в виде какого звена пищевой цепи целесообразно получать хозяйственно-ценную продукцию, необходимо будет планировать и состав уловов, а "ели окажется целесообразным базировать уловы на хищной рыбе, то планировать и такой режим эксплуатации хищной рыбы, который обеспечивал бы получение максимума продукции. Если же будет признано целесообразным получать основную продукцию в виде мирных рыб, то численность хищной рыбы должна быть тем или иным способом ограничена.
Однако в большинстве водоемов та или иная часть кормовых ресурсов как первого, так и второго звеньев трофических цепей не может быть использована ни молодью, ни взрослыми промысловыми рыбами. В Амуре, например, значительная часть русловых личинок Chironomidae используется только мелкими пескарями, которые в свою очередь уже служат пищей промысловым хищным рыбам. Поэтому во многих водоемах определенная часть вылова всегда должна планироваться и в виде хищных рыб. Планирование видового состава уловов должно основываться на знании пищевых отношений в водоеме еще и потому, что чрезмерная интенсификация промысла одного вида часто может приводить к увеличению численности других, обычно менее ценных видов (часто принадлежащих к другим фаунистическим комплексам), популяции которых развиваются за счет кормов, недоиспользуемых интенсивно облавливаемыми популяциями промысловых видов. Такие примеры известны в отношении и морских, и пресноводных рыб (Larkin, 1963).
Таким образом, при планировании интенсификации вылова того или иного промыслового вида одновременно надо увеличивать вылов и малоценных видов, потребляющих те же корма, что и ценный промысловый вид, т. е. необходимо планировать эксплуатацию биологических ресурсов экосистемы водоема как единого целого.
Очень важно также, чтобы промысел изымал промысловую рыбу в основном уже после того, как популяция в данном сезоне использовала свою кормовую базу, т. е. закончила основной период нагула. В отношении полупроходных рыб наших южных морей - леща, сазана, судака и других - 'это должна быть осень, в отношении лососей - начало нерестовой миграции и т. д.
При расчете величины вылова промысловых видов рыб в водоеме приходится учитывать происходящие в нем в разные годы изменения кормовой базы. Если кормовая база обеспечивает тому или иному виду в данном году больший прирост биомассы популяции, то величину его вылова можно увеличить. При этом надо иметь в виду, что максимальная продуктивность популяции достигается не при максимальном или минимальном индивидуальном темпе роста особей, а при средних величинах темпа роста. При самом быстром росте, т. е. в разреженных посадках, корма используются менее полно и эффективно, чем в посадках средней плотности. В переуплотненных посадках соотношение поддерживающего и продуцирующего кормов меняется в пользу первого и общая продуктивность популяции также снижается. Примеры этого были приведены выше. Надо при этом также иметь в виду, что "ответ" популяции на изменение кормовой базы у рыб с коротким жизненным циклом будет быстрее, чем у рыб с длительным жизненным циклом и поздней половозрелостью.
Наконец, очень важный метод более полного освоения кормовой базы водоема - это соответствующий подбор возрастной структуры стада, при которой полнее всего осваиваются корма. У большинства видов рыб, особенно в умеренных и низких широтах, и у достигших половой зрелости рыб состав пищи и нагульные площади рыб разного размера и возраста различны. Следовательно, интенсивно облавливая старшие возрастные группы и сокращая их численность, мы оставляем недоиспользованной часть кормовых ресурсов, которыми не питаются особи младших возрастных групп, а иногда остаются без должного использования и кормовые ресурсы отдельных районов, где кормятся только старшие возрастные группы. Так, например, повторно нерестующие особи атлантическо-скандинавских сельдей осваивают для нагула более западные и северные районы Атлантики (район Шпицбергена), чем впервые нерестующие особи (район Баренцева моря). Сокращение численности старших возрастных групп сельди приводит к недоиспользованию кормовой базы этих районов (Марти, 1956).
Важно, как это отмечает Марти (1956), что разница в распределении связана с размерами в большей степени, чем с возрастом рыбы. Быстро растущие особи младших возрастов, близкие по размерам к рыбам старших возрастов, мигрируют на нагул вместе с последними.
Лещ Аральского моря старших возрастов осваивает более глубокие части Аральского моря, занятые грунтами в виде серого ила с высокой биомассой мотылей, которые совсем не осваиваются младшими возрастными группами леща (рис. 112) (Бервальд, 1961). Различия в составе пищи и в местах нагула отмечены также у леща Каспия и леща Азовского моря (Бердичевский, 1964). У арктической трески Gadus morhua morhua L.
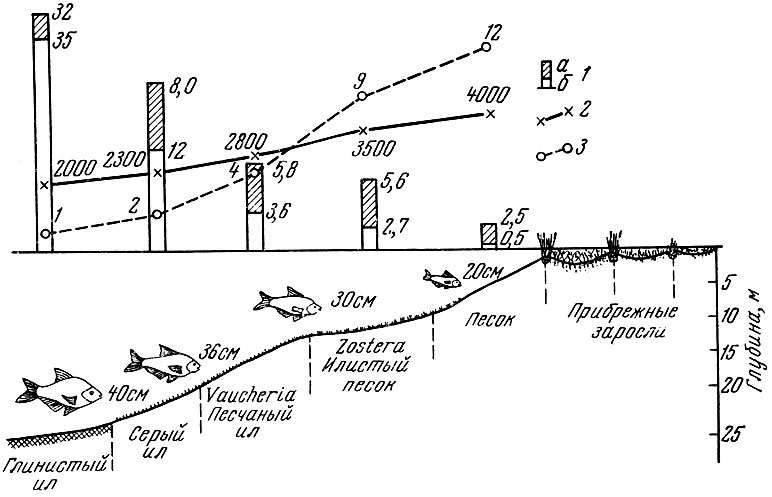
Рис. 112. Распределение средней биомассы бентоса в июне 1951 г. (в г/м2) (2), суммы градусо-дней в году (2) и продолжительность пребывания рыбы на биотопах (в месяцах) (3) (по Бервальду, 1961): а - моллюски без раковин; б - мотыль. Числа возле контуров рыб - средний размер нагуливающегося леща (в см)
самые старые и повторно нерестующие особи держатся в основной массе западнее и севернее, чем впервые нерестующие. Более крупная треска также менее привязана ко дну, чем впервые нерестующая. Различен, как известно, и состав пищи трески разных возрастов. Особи старших возрастов речной камбалы Pleuronectes flesus L. в Балтийском море питаются крупными моллюсками, держащимися глубже, чем моллюски, которыми кормятся более мелкие и молодые особи камбал (Желтенкова, 1954). Подобных примеров различий в составе потребляемых кормов и местах нагула особей разного размера и возраста в пределах популяции одного и того же вида можно привести очень много (Бердичевский, 1964).
Практически у большинства основных промысловых рыб в водоемах всех географических широт и после перехода на основной для вида тип питания имеются значительные расхождения в составе пищи и местах нагула особей разных размерных групп. Некоторые же виды на разных этапах онтогенеза входят даже в разные экосистемы.
Конечно, у рыб старших возрастных групп эффективность использования кормов ("оплата корма") обычно ниже, чем у более молодых. Однако из этого вовсе не вытекает необходимость перестраивать популяции промысловых рыб так, чтобы они состояли почти исключительно из впервые нерестующих рыб, тем более что состав пищи повторно нерестующих рыб, как указывалось, обычно отличен от состава пищи впервые нерестующих, и влияние численности поколений рыб старшего возраста обычно почти не отражается на росте особей во впервые созревающих поколениях (Поляков, 1962).
К сожалению, мы еще очень плохо знаем, как у разных видов промысловых рыб меняется с возрастом "оплата корма" - соотношение поддерживающего и продуцирующего кормов, как меняется это соотношение в зависимости от количества потребляемого корма и режима потребления. Исследования по этим разделам физиологии питания и роста необходимо интенсивно развивать. Конечно, наличие большого количества старых рыб в популяции, у которых снизилась или частично утрачена способность воспроизводства, с точки зрения продукционных возможностей популяции обычно нецелесообразно. Однако надо иметь в виду, что у рыб старших возрастов процесс старения, качество продуцируемых половых продуктов, периодичность размножения очень тесно связаны с обеспеченностью пищей. Если кормов много, то регулирование рыболовства путем резкого сокращения численности рыб старших возрастов может быть и нецелесообразным; в случае же низкой обеспеченности пищей наличие старых рыб в популяции может значительно снижать ее продуктивность.
При планировании возрастного состава уловов необходимо знать возрастную структуру впервые созревающих особей, ее изменения в связи с изменением условий жизни и периодичность нереста. Надо знать и характер связи возраста полового созревания с продолжительностью жизни, а также возрастную структуру стареющих особей.
Таким образом, характер использования популяцией кормовой базы служит важным критерием для расчета режима ее эксплуатации.
Существеннейшими показателями, по которым должен строиться расчет режима эксплуатации стад промысловых рыб, служат как нарастание массы тела у особей разного возраста, так и общая величина биомассы отдельных возрастных групп, являющаяся результирующей процесса весового роста особей в данном поколении. У большинства видов рыб нерестовых популяций второго и третьего типов максимальные весовые приросты приходятся на особей среднего возраста, обычно повторно нерестующих. У леща, например, - это 6-7 лет, у судака - 4-5 лет и т. д. (Бервальд, 1956; Бердичевский, 1961, 1964).
Значительно сложнее расчет убыли популяции. Непосредственное определение убыли по возрастному составу стада даже на основе средних многолетних данных может привести к очень значительным ошибкам, особенно в отношении видов рыб со значительными флюктуациями. Поэтому расчеты носят, естественно, лишь ориентировочный характер.
У относительно крупных рыб наших южных водоемов, таких, как лещ и сазан, не говоря уже об осетровых, естественная смертность в среднем возрасте, видимо, как правило, невелика, Несколько больше, но также очень незначительна она у таких рыб, как треска, морской окунь и др. Если принять даже завышенные нормы естественной смертности, то и тогда перенос основного воздействия промысла на более старшие возрастные группы дает очень значительное увеличение выхода рыбной продукции, а так как крупная рыба обычно ценнее как пищевой продукт, то при этом улучшается и качество улова.
У арктической трески при перенесении основного вылова с особей в возрасте 7 + на особей в возрасте 9 + в результате естественной смертности (по данным рабочей группы по треске Международного совета по изучению моря*) теряется около 20% особей, а прирост биомассы получается около 125%. У плотвы соответственно при вылове в среднем возрасте 4 года теряется по сравнению с выловом в возрасте 5 лет около трети улова. У среднерусской плотвы естественная смертность в возрасте от 4 + до 6 + невелика.
* (Сб. научн. информации ВНИРО, 1959, № 14.)
Как видно из табл 105, весь улов аральского леща в перспективе мог быть на 176% больше, чем в 1939 г., при вылове одного и того же количества особей, но в более старом возрасте.
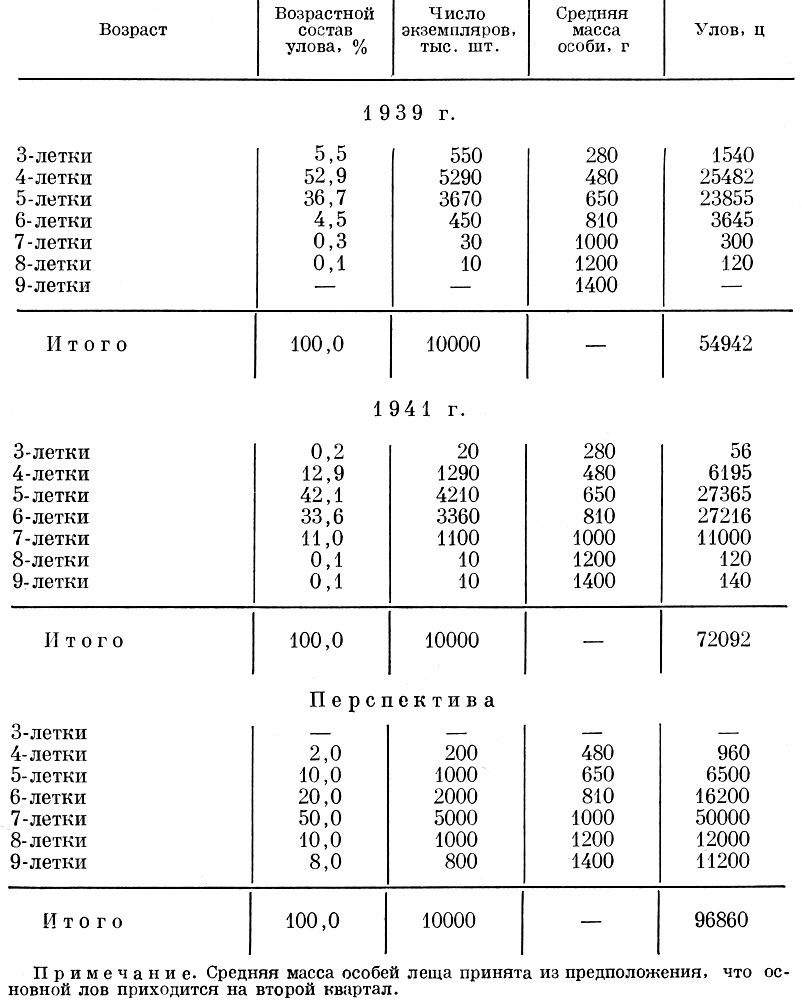
Таблица 105. Зависимость величины вылова от возрастного состава улова аральского леща (по Бервальду, 1956; расчет ведется условно на 10 млн. экз. леща)
Много примеров подобного рода потерь при нерациональной эксплуатации приведено в отношении рыб наших южных морей Л. С. Бердичевским (1961, 1964).
Естественно, что повышение биомассы вылова популяции путем регулирования возрастного состава уловов дает значительный эффект лишь в отношении рыб с относительно продолжительной жизнью и малой смертностью в средних возрастах. У рыб с коротким жизненным циклом, у которых остаток в нерестовом стаде составляет малый процент от пополнения, перенесение вылова на более старшие возрасты, как правило, дает малый эффект. Наибольший улов у таких рыб достигается при облове впервые нерестующих, достигших половозрелости рыб. Роль остатка в улове может быть минимальной.
Важное значение для повышения продуктивности популяций имеет правильная сезонная дислокация уловов. Выше уже говорилось, что для более полного использования кормовой базы рыбу желательно вылавливать по окончании сезона нагула, когда она максимально использовала свою кормовую базу. Естественная смертность у промысловых рыб с относительно длительным жизненным циклом за период от начала до конца нагула обычно невелика.
Конечно, не у всех рыб и не во всех географических широтах наблюдается значительная сезонная динамика интенсивности питания, а следовательно, жирности и упитанности. Невелика сезонная динамика интенсивности питания и жирности, например, у щуки. Слабо выражена она у ряда тропических рыб; жирность и упитанность многих из них также существенно не меняется в разные сезоны.
В умеренных и частично высоких широтах в отличие от экваториальных и тропических вод у популяций животных и особенно у рыб резко выражена сезонность концентрации энергетических ресурсов. Как правило, максимальной биомассы популяция рыб достигает к осени, когда заканчивается сезон ее нагула. К этому же времени в популяции концентрируется и наибольшее количество жиров. Так, треска в водах Исландии содержит весной на 1в% меньше жира и белка, чем осенью. Печень у трески в Баренцевом море осенью весит намного (до 70%) выше, чем весной. Беломорская корюшка содержит осенью на 21% больше энергетических ресурсов, чем весной. Только во время нереста самки корюшки теряют около 60% накопленных в организме жиров, из которых около 50% концентрируется в икре и идет на обеспечение желточного питания потомства, а 10% расходуется на покрытие трат энергии во время размножения. Аральский лещ осенью содержит на 50-60% больше жира, чем весной, и одновозрастные особи половозрелого леща весят осенью на 12-20% больше (Бервальд, 1956, 1961). Близкая картина наблюдается и у сельди; основной ее улов, правда, несколько меньший, чем трески, приходится на весеннее время, и при этом теряется около 15-20% питательных веществ.
Таким образом, только путем перенесения основного сезона лова трески, корюшки, леща и других рыб с весны на осень общий улов может быть значительно увеличен и намного возрастет выход жира. В настоящее время основная добыча рыбы часто осуществляется в период размножения, когда рыба оказывается более всего доступной для облова. Такой лов оказывается экономически выгоднее, чем добыча товарно более ценной рыбы в другое время года, но труднее поддающейся облову. Ввиду этого перед специалистами по технике лова и ихтиологами, занимающимися изучением поведения рыб, возникает очень важная задача - разработать такие способы добычи, которые делали бы рентабельным вылов рыбы, когда она как пищевой продукт находится в наиболее ценном состоянии.
Достижение максимальной продуктивности эксплуатируемых популяций рыб требует также правильной дислокации добычи в пространстве. Современная концентрация вылова многих рыб (например, воблы, сахалинской сельди и др.) на нерестилищах часто нарушает ход естественного воспроизводства. Во многих случаях значительный прилов молоди объясняется не только мелкой ячеей в орудиях лова, но и добычей в таких местах и в такое время, когда в районах лова скапливается большое количество молоди (например, молодь пикши на Канинской банке). Правильная дислокация добычи рыбы в пространстве часто позволяет обеспечить необходимый возрастной состав уловов и без регулирования размеров ячеи. Это особенно важно в отношении тех видов рыб, у которых регулирование размеров особей в уловах при помощи ячеи бывает мало эффективным (например, сельдь).
Таким образом, продуктивность стад промысловых рыб может быть повышена путем: 1) планирования правильного возрастного состава уловов; 2) правильного сезонного графика лова; 3) правильной дислокации добычи в пространстве; 4) установления определенной нормы вылова. Естественно, что режим эксплуатации популяций разных видов будет иметь ряд общих черт, но вместе с тем он будет характеризоваться и своей спецификой. Вылов всех видов рыб, за очень редкими исключениями, должен базироваться на созревающих или уже достигших половой зрелости особях. В отношении рыб, нерестующих 1 раз в жизни (нерестовая популяция первого типа), или рыб с очень незначительным процентом повторно нерестующих особей установление минимальной меры допускаемых к вылову особей при облове нерестовых популяций, мигрирующих к местам размножения, безусловно нецелесообразно. Это относится, например, к дальневосточным и атлантическому лососям, к омулю, большинству проходных сельдей и ряду других рыб. Регулированием размерного и возрастного состава особей в нерестовом стаде можно добиться наибольшего эффекта у рыб с длительным жизненным циклом, у которых повторно нерестующие рыбы составляют значительный процент от всей популяции.
В низких широтах, где жизненный цикл рыб обычно короче, регулирование возрастного состава нерестового стада у многих промысловых рыб, как правило, не может дать значительного увеличения улова.
В тропиках и экваториальной зоне основное внимание в отношении большинства важных промысловых видов должно быть направлено на защиту от вылова неполовозрелой части стада. Это в первую очередь относится к большинству окунеобразных, кефалеобразных и камбал.
Расчет желательного возрастного состава уловов должен базироваться на следующих показателях:
1) весовом приросте (индивидуальном) рыбы разных возрастов;
2) изменении биомассы каждой возрастной группы (включая анализ естественной смертности на разных возрастах);
3) спектре питания каждой возрастной группы и характере использования популяцией кормовой базы в целом;
4) "оплате корма" разными возрастными группами и при разных соотношениях концентраций корма и кормящейся рыбы;
5) качестве половых продуктов, продуцируемых особями разного возраста;
6) темпе роста особей разных поколений.
Общий расчет необходимой структуры изымаемой популяции несколько варьирует из года в год в зависимости от состояния кормовой базы и численности отдельных поколений в пополнении (последнее корректируется через изменения темпа роста).
В отношении даже основных промысловых рыб мы еще не имеем пока полного комплекса необходимых данных для расчета оптимального возрастного состава уловов и приходится базироваться хотя бы на минимальном наборе необходимых сведений.
Неправильно ориентировать вылов таким образом, чтобы уловы большинства промысловых рыб состояли почти исключительно из быстрорастущих впервые нерестующих особей (Терещенко, 1913; Чугунов, 1928,идр.). При такой эксплуатации стад, как правильно показано Л. С. Бердичевским (1964), общий выход рыбного мяса оказывается ниже, чем при эксплуатации многовозрастной популяции. Кроме того, потомство, получаемое от впервые нерестующих рыб, обычно обладает меньшим запасом желтка и в изменчивых условиях часто оказывается менее жизнестойким, чем потомство, получаемое от повторно нерестующих рыб. Как указывалось выше, очень важным элементом, обеспечивающим высокую продуктивность стада, является установление нормы вылова (Никольский, 1950, 1972).
Однако я считаю совершенно неправильным противопоставление "контингента вылова" правилам рыболовства (Баранов, 1918, 1925). Выше было показано, как значительно может меняться возможный вылов в зависимости от того, в каком возрасте и в какое время года добыта рыба. Поэтому научно обоснованная, допустимая норма вылова должна сочетаться с определенной регламентацией режима эксплуатации, а не противопоставляться ей. Нормирование и дислокация вылова по районам и сезонам должны входить в общую систему мероприятий, слагающих единую систему рациональной эксплуатации стад промысловых рыб.
Для разработки правильной сезонной и пространственной дислокации добычи необходимо опираться на данные о: 1) сезонной динамике биологических показателей у разных возрастных групп в популяциях промысловых рыб (жирность, упитанность, прирост массы, ход созревания половых продуктов и др.); 2) дислокации в пространстве рыб разного возраста и биологического состояния и закономерностей их перемещений; 3) закономерностях, которым подчиняются образование концентраций промысловых рыб, места и время их образования.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'