Влияние вылова на стадо промысловой рыбы
Вылов оказывает на стадо промысловой рыбы весьма разнообразное влияние. Изымая часть стада, вылов через разреживание популяции повышает обеспеченность пищей оставшейся части стада, что связано с изменением темпа роста особей, возраста достижения половой зрелости и предельного возраста. Селективный вылов, изымая из популяции ту или иную часть, сказывается на изменении структуры популяции, а тем самым и на ее воспроизводительных свойствах.
Изымая часть стада, вылов неминуемо изменяет интенсивность, а иногда и характер воздействия стада рыб на ее кормовую базу, создавая в ряде случаев благоприятные условия для питания других видов рыб, потребляющих корма, сходные с кормами вида рыбы, являющейся объектом рыболовства.
Разреживая стадо хищной рыбы, вылов меняет интенсивность и характер ее воздействия на стадо мирной рыбы. В отношении стайных рыб, у которых стая является защитным приспособлением от хищников, вылов, нарушая структуру стаи, часто может делать этих рыб более доступными для воздействия хищников. Наконец, в ряде случаев вылов меняет паразитарные связи промысловой рыбы.
По своему действию на популяцию вылов напоминает действие хищников, и реакция популяции на вылов во многом сходна с реакцией на воздействие хищников. Разница обычно заключается в том, что вылов действует главным образом на половозрелую часть стада, а хищники на неполовозрелую.
У интенсивно облавливаемых популяций промысловых рыб темп роста выше и популяционная плодовитость больше. Однако у всех популяций рыб повышение интенсивности рыболовства вызывает нарастание темпа роста и плодовитости лишь до определенных пределов, после чего, если интенсивность рыболовства превзойдет определенную, специфичную для каждого вида величину, регуляторные механизмы популяции нарушаются и она перестает реагировать на дальнейшее разреживание стада, что является очень серьезным сигналом перелова. Виды рыб с коротким жизненным циклом, ранней половой зрелостью, большим пополнением по отношению к остатку, приспособленные к интенсивному воздействию хищников на половозрелую часть популяции, допускают и больший процент изъятия половозрелой популяции промыслом. Рыбы же с многовозрастной структурой популяции, поздним половым созреванием, с относительно малым пополнением по отношению к остатку, приспособленные к относительно малой смертности от хищников на старших возрастах, допускают и относительно малый процент изъятия по отношению ко всей половозрелой части стада.
Таким образом, воспроизводство популяции не нарушается при определенной (для каждого вида) интенсивности вылова. Это происходит тогда, когда вылов изымает такую часть стада, к изъятию которой популяция приспособлена (выедание хищниками), когда вылов может быть компенсирован регуляторными механизмами популяции, т. е. когда вылов как бы является элементом среды вида. При подобной интенсивности вылова и, конечно, если не будут нарушены нерест и условия развития молоди, популяция сможет существовать многие годы, ежегодно обеспечивая определенную величину вылова.
У каждого вида рыб в процессе его становления и приспособления к другим видам данного комплекса вырабатывается система приспособлений, защищающих его от хищников, и система регуляторных механизмов, компенсирующих определенную смертность, т. е. в процессе исторического развития вид приспосабливается к определенной интенсивности смертности и определенной ее амплитуде. При этом вид приспосабливается к определенной интенсивности смертности в каждой возрастной группе. Вылов является новым элементом среды, входящим в жизнь уже сформировавшегося вида.
Виды, хорошо защищенные от хищников структурными защитными приспособлениями (шипы, колючки и др.) и системой поведенческих реакций, часто оказываются особенно легко доступными для добычи. Всякое приспособление конкретно (Васнецов, 1938, 1953). Например, шипы и колючки, являющиеся защитными по отношению к хищным рыбам, делают рыбу более уязвимой в отношении сетных орудий рыболовства. Как показал Ю. Ю. Марти (Marty, 1961), очень многие защитные приспособления от хищников являются вредными для рыбы при воздействии рыболовства (табл. 88).

Таблица 88. Относительность защитных приспособлений у рыб (по Marty, 1961, с изменениями)
В то же время хорошо защищенные от хищников виды обычно характеризуются сравнительно низкой воспроизводительной способностью, и их интенсивный вылов часто не может быть компенсирован. Если рыбы защищены от хищников системой безусловных и условных рефлексов, то защитных от орудия лова безусловных рефлексов у рыб нет, а выработка условных рефлексов на орудия лова весьма затруднена, так как для того, чтобы выработался условный рефлекс, рыба должна несколько раз вырываться из орудий лова.
Правда, как показали опыты Д. С. Николаева, у рыб удается вырабатывать условные рефлексы на колющие орудия (крючки), причем у разных видов пресноводных рыб скорость выработки условного рефлекса и его стойкость оказываются весьма различными. Отмечено, что у рыб вырабатывается условный рефлекс на подводного охотника, и они держатся от него на дистанцию выстрела (Мантейфель, 1970).
Таким образом, сравнивая вылов с воздействием хищников, при организации эксплуатации стад промысловых рыб и их воспроизводства мы должны учитывать и то общее, что имеется в характере их воздействия и существующие различия. И то и другое надо максимально использовать для обеспечения высокой продуктивности популяций промысловых рыб.
Как видно из изложенного, общий мировой улов продолжает расти. Конечно, этот рост не может быть беспредельным. При современном экстенсивном, очень слабо регулируемом морском рыбном хозяйстве, видимо, максимальный возможный вылов может быть довольно скоро достигнут. И сейчас уже наряду с общим ростом улова в добыче отдельных видов рыб наблюдается стабилизация, а в ряде случаев даже довольно быстрое снижение уловов. Так, например, уловы акулообразных за последние годы несколько уменьшились; уловы некоторых особенно интенсивно промышляемых видов акулообразных настолько упали, что промысел свелся почти на нет. Весьма поучителен в этом отношении промысел в первую очередь суповой акулы Galeorhinus zyopterus Jordan a. Gilbert по Тихоокеанскому побережью США (Ripley, 1946). Общий улов акул в Калифорнии (рис.70) с 1930 по 1937 г. находился на одном уровне и колебался от 471 тыс. англ. фунтов в 1933 г. (английский фунт равен 453,592 г) до 914 тыс. англ. фунтов в 1937 г. В 1938 г. улов увеличился более чем до 7,5 млн. англ. фунтов. В 1939 г. он достиг максимальной величины - 9228 тыс. англ. фунтов, и с этого года начал быстро падать. В 1944 г. он составлял уже только немногим более 2,5 млн. англ. фунтов, а в 1959 г. упал до 602 тыс. фунтов, т. е. дошел до уровня 1933-1937 гг. Еще стремительнее уменьшались уловы суповой акулы: примерно с 5 млн. в 1941 г. до 600 тыс. англ. фунтов уже в 1944 г., т. е. запасы были подорваны за какие-нибудь 6-7 лет.
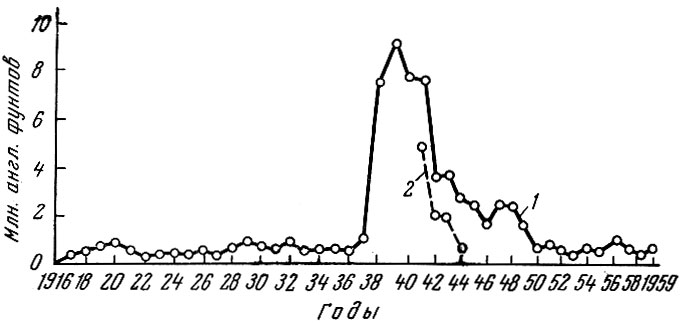
Рис. 70. Уловы акул в Калифорнии (по Ripley, 1946, с добавлениями): 1 - общий улов; 2 - уловы суповой акулы
Сильно снизился улов осетровых рыб. Несмотря на значительную интенсификацию рыболовства, уловы лососей не только не возросли, но в основных районах рыболовства даже снизились (Cooley, 1963). После второй мировой войны подъем уловов очень скоро сменился снижением.
Очень характерен в этом отношении пример австралийского промысла тигрового плоскоголова Neoplatycephalus macrodon (Ogilby) с малых тральщиков. Этот промысел начал усиленно развиваться в середине второй мировой войны и достиг максимума к концу войны, когда и общий улов, и улов на рыболовное усилие оказались наибольшими (рис. 71). Затем улов начал быстро падать, и значение промысла этого вида намного снизилось.
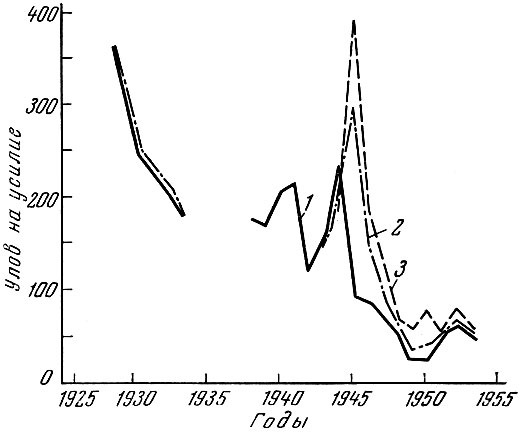
Рис. 71. Уловы тигрового плоскоголова (по Fairbridge, 1952): 1 - паровые траулеры; 2 - датские сейнеры; 3 - паровые траулеры и датские сейнеры
Не менее характерна история промысла палтуса Hippoglossus hippo-glossus stenolepis Schmidt по Тихоокеанскому побережью США, подробно исследованная Томпсоном (Thompson, 1950, и др.) и его сотрудниками. Нерегулируемое рыболовство привело к резкому падению сначала уловов на рыболовное усилие, а затем и общего улова. Только после осуществления ряда мероприятий по упорядочению рыболовства уловы снова повысились (рис. 72). Это изменение уловов связано с изменением структуры облавливаемой популяции: изменились темп роста рыб, время полового созревания, возрастной состав и другие биологические показатели.
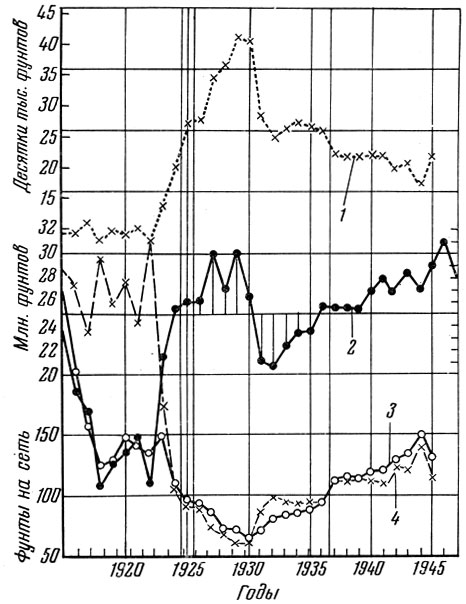
Рис. 72. Уловы палтуса (по Thompson, 1950): 1 - число порядков сетей; 2 - годовой улов; 3 - улов на порядок сетей; 4 - нормальный улов на порядок
Сходные изменения (табл. 89) произошли в стаде камбал залива Петра Великого в результате развития интенсивного промысла с малых тральщиков в местах зимнего скопления камбал (Моисеев, 1946).

Таблица 89. Уловы камбал тральщиками в районе зимнего скопления в заливе петра великого
Колебания величины уловов были связаны также с изменением среднего размера камбал в промысловых уловах, времени их полового созревания и возрастного состава стад.
Существенно изменился также видовой состав уловов камбал. Так, в 1930/31 г. один из наиболее ценных промысловых видов Limanda aspera (Pall) составила 60-70% всего улова камбал, а в 1935/36 г. только несколько (единицы) процентов. Уловы же Сleisthenes her zensteini (Schmidt) возросли с 10-15% в 1930/31 г. до 35% в 1935/36 г. (Моисеев, 1946).
Сходная картина наблюдалась с уловами камбал по американскому побережью Атлантического океана (Merriman a. Warfel, 1948). Интенсивный промысел "зимней камбалы" начался во втором десятилетии нашего столетия и достиг максимума в 1930 г., когда улов камбал, главным образом Рseudopleuronectes americanus (Wall.) и Limanda ferruginea (Storeг), составил 11 млн. англ. фунтов. На этом уровне он держался некоторое время благодаря увеличению флота и радиуса его действия, а затем последовало снижение уловов в 1943-1945 гг., когда интенсивность промысла этих прибрежных камбал резко возросла (табл. 90).
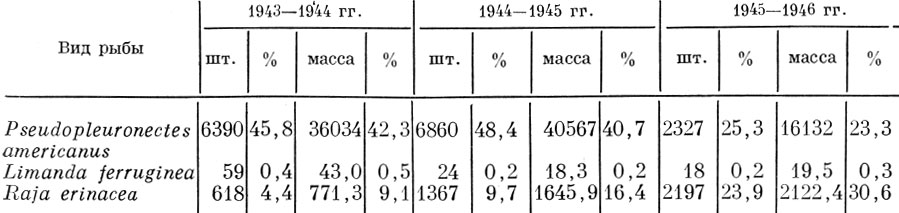
Таблица 90. Улов (в шт.) и биомасса улова двух видов камбал (улов на час траления) и малого ската у берегов южной новой англии (по Merriman a. Warfel, 1948)
Несколько более сложная картина наблюдалась с рыбами, характеризующимися близким типом динамики стада, в европейских водах. Здесь дважды в результате запуска рыболовства в период первой и второй мировых войн резко снижалась интенсивность рыболовства, что приводило к восстановлению стада. При этом происходили соответствующие изменения и в структуре популяции, и в темпе роста рыб, ее слагающих. Наиболее полные материалы в этом отношении имеются о морской камбале Platessa platessa L. Северного моря (Thursby-Pelham, 1939; Buckmann, 1939; Jensen, 1948, и др.). Общий улов морской камбалы в Северном море колебался от 25 до 72 тыс. т. Уловы в период как первой, так и второй мировой войн (рис. 73) снижались, а затем резко возрастали после окончания войны (табл. 91).
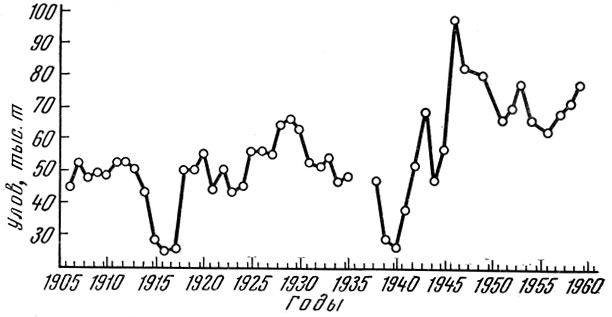
Рис. 73. Уловы морской камбалы в Северном море (по Margets a, Holt, 1948 и по данным Bulletin Statistique)
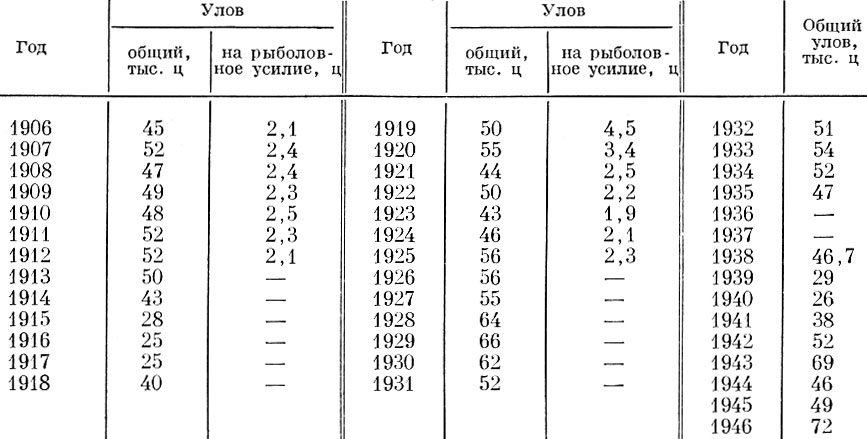
Таблица 91. Уловы морской камбалы в северном море (по данным 1948 г.)
Снижение интенсивности рыболовства и падение почти вдвое уловов за время первой мировой войны привело к восстановлению стада камбалы, уловы на рыболовное усилие в послевоенное время возросли более чем в 2 раза. Сходная, но менее резко выраженная картина получилась и в результате запуска во время второй мировой войны (Margets a. Holt, 1948).
Сравнение темпа роста камбалы до и после второй мировой войны показало (рис. 74 и 75), что в большинстве сравниваемых проб темп роста и средние размеры рыб соответствующих возрастных групп оказались более высокими в предвоенных уловах.
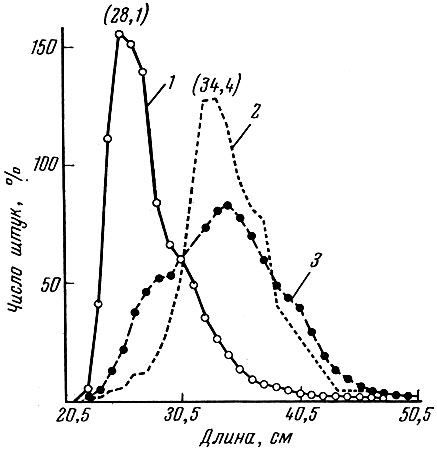
Рис.74. Размерный состав морской камбалы (в %) в июле 1938 (1), 1945 (2) и 1946 (3) гг., доставляемой в Ловестофт (по Margets a. Holt, 1948)
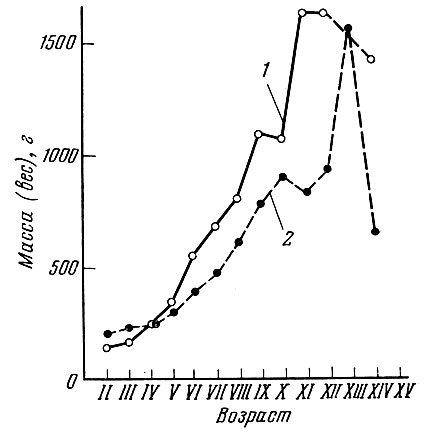
Рис. 75. Средняя масса каждой возрастной группы из промысловых уловов морской камбалы Северного моря в декабре 1938 (1) и 1946 (2) гг. (по Margets a. Holt, 1948)
Изменился возрастной состав уловов. Как видно на рис. 30, средний возраст камбалы в уловах увеличился, причем среди рыб улова 1945 г. совсем не было зрелых особей двух- и трехлетнего возраста.
Сильно изменился и размер рыб в уловах. Средние размеры камбалы в уловах 1945 и 1946 гг. оказались много выше, чем в 1938 г. (см. рис. 74). Однако последующее увеличение интенсивности рыболовства в послевоенные годы снова привело к омоложению стада и ускорению роста рыб, связанному с более ранним достижением половозрелости. Правда, принятые меры по увеличению размеров ячеи в орудиях лова обеспечили некоторое сокращение в уловах количества маломерных рыб (Gulland, 1972).
И в южных морях, омывающих Европу, на ряде видов также можно проследить характер изменений, вызываемых воздействием рыболовства. В Азовском море, например, как это хорошо показал Н. Л. Чугунов (1927), в результате запуска рыболовства в период первой мировой и гражданской войн 1914-1920 гг. средние размеры и возраст осетровых в уловах значительно увеличились, а темп роста замедлился. Последующая интенсификация промысла привела к уменьшению средних размеров и возраста осетровых и усилению темпа их роста.
Мы рассмотрели воздействие рыболовства на те виды рыб, у которых урожайность поколений по годам меняется сравнительно незначительно и мощность поколений отдельных лет различается обычно не более чем в 10 раз. Попытаемся теперь рассмотреть, как отражается рыболовство на стадах тех рыб, у которых наблюдаются значительные колебания урожайности отдельных поколений (флюктуации). У арктической трески, основные нерестилища которой расположены по северо-западному побережью
Норвегии и промыслом которой на нагульных площадях сейчас заняты очень многие страны, до 1957 г. не наблюдалось закономерной "длящейся" тенденции к снижению общего улова (рис. 76).
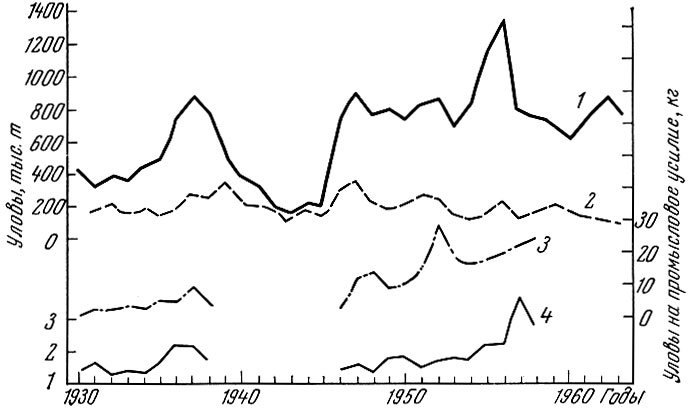
Рис. 76. Уловы арктической трески (в тыс. т и на промысловое усилие) в 1930 - 1960 гг. (по данным рабочей группы по арктическому рыболовству. 'Научная информация', № 14, 1959): 1 - в трех промысловых районах; 2 - у побережья Норвегии; 3 - общее промысловое усилие; 4 - общее промысловое усилие в районе Лофотен (в норвежских единицах)
В отличие от рыб предыдущей группы, уловы трески в разные годы очень резко колеблются, что несомненно связано с резкими колебаниями урожайности поколений. Однако если до 1914 г., как это отмечает Йорт (Hjort, 1914), норвежские уловы трески на рыболовное усилие тоже росли, то в 30-е годы они несколько упали, хотя общий улов к 1939 г. даже поднялся. В послевоенные годы наблюдался значительный подъем уловов. Максимальный улов трески в северной части Атлантического океана был в 1956 г. Начиная же с 1957 г. наблюдается общее падение уловов. Как хорошо показано Роллефсеном (Rollefsen, 1954), изменения численности стада трески связаны с изменением темпа роста и временем полового созревания. Процент впервые нерестившихся рыб среди особей старших возрастов в период запуска последней войны сильно понизился, а в послевоенный период возрос. В конце 60-х и начале 70-х годов уловы трески продолжали падать, что связано как с чрезмерной интенсивностью промысла, так и с низкой урожайностью поколений ряда лет.
Несколько иная, чем с треской, картина наблюдалась с пикшей Melanogrammus aeglefinus(L.) (рис. 77). Уловы ее в исландских водах перед первой мировой войной держались примерно на одном и том же уровне, но уловы на рыболовное усилие начиная с 1911 г. стали сильно снижаться. После первой мировой войны произошло резкое повышение общих уловов, уловы же на рыболовное усилие сразу стали резко снижаться. Общие уловы пикши продержались на высоком уровне до 1929 г., после чего также начали быстро падать. Если рассмотреть уловы всех европейских стран во всем Северном море (Raitt, 1939), то картина, хотя и окажется несколько более сглаженной, но тоже достаточно четкой.
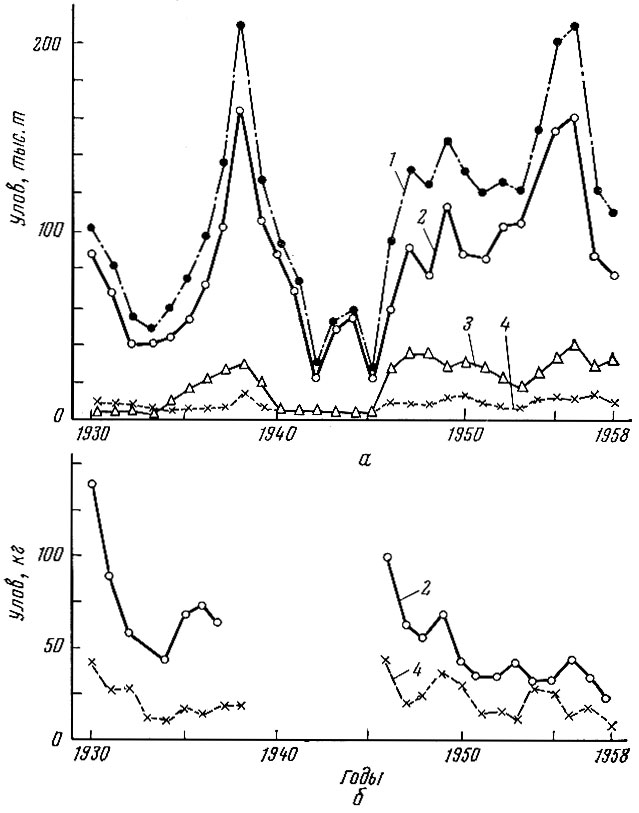
Рис. 77. Общие уловы арктической пикши по районам (а) и улов на единицу промыслового усилия (б) за 1930- 1958 гг. (по материалам рабочей группы): 1 - по всем районам; 2 - в Баренцевом море; 3 - у побережья Норвегии; 4- , у о-вов Медвежий и Шпицберген
| Год | Улов, тыс. кг |
|---|---|
| 1919 | 204012 |
| 1920 | 210012 |
| 1921 | 171291 |
| 1922 | 165330 |
| 1923 | 124129 |
| 1924 | 106617 |
| 1925 | 147083 |
| 1926 | 130125 |
| 1927 | 125223 |
| 1928 | 114703 |
| 1929 | 107744 |
| 1930 | 129378 |
| 1931 | 109893 |
| 1932 | 97913 |
| 1933 | 108032 |
| 1934 | 85994 |
| 1935 | 67328 |
| 1936 | 56609 |
В послевоенные годы уловы пикши в Северном море опять резко возросли.
Улов на рыболовное усилие (улов за 100 ч работы парового тральщика) в 1946 г. увеличился по сравнению с 1938 г. в 4 раза (в 1946 г. - 34, в 1938 - 12 cwt) (Margets a. Holt, 1948). Однако это увеличение уловов довольно скоро сменилось опять понижением.
Интенсификация вылова пикши в предвоенные годы привела к разреживанию ее популяции и ускорению роста особей; это изменение темпа роста было связано с более ранним созреванием и омоложением возрастного состава нерестового стада (рис. 78). Наоборот, период запуска во время войны привел к постарению нерестовой популяции (Parrish, 1948).
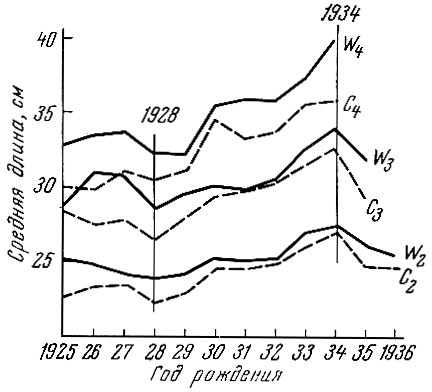
Рис. 78. Скорость роста пикши на втором, третьем и четвертом годах жизни поколений 1925-1936 гг. в западной (W) и центральной (С) частях Северного моря (по Raitt, 1939)
С середины 50-х годов наблюдалось снижение уловов и ускорение роста рыб.
Некоторый подъем уловов североморской пикши в последние годы связан с появлением высокоурожайного поколения 1965 г.
Более сложная картина наблюдается в отношении сельдей, населяющих Северо-Восточную Атлантику.
До недавнего времени большинство исследователей считало, что существующий промысел не полностью использует стада атлантической сельди и влияние вылова слабо сказывается как на структуре популяции, так и на общей численности стад. Этим, в частности, объясняется тот факт, что в Международной конвенции, заключенной в 1946 г. для регулирования рыболовства в Северо-Восточной Атлантике, предусмотренные этой конвенцией охранные меры распространялись только на тресковых и камбаловых рыб, промысел же сельди не регулировался. Основным критерием для такого вывода было значительное возрастание уловов атлантическо-скандинавских сельдей и сельдей Северного моря. Однако за последние годы как общие уловы ряда стад атлантической сельди, обладающих различной численностью, так и уловы на рыболовное усилие снижаются. Колебания уловов сельди были известны с давних времен, но снижения уловов, как правило, не носили длительного характера. В настоящее время в отношении некоторых стад наблюдается довольно длительная депрессия уловов. Это свидетельствует о том, что интенсивность промысла этих стад достигла такой величины, которая не может быть компенсирована пополнением при обычных (средних) условиях воспроизводства. Это приводит к прогрессивному снижению численности стада (Никольский и Белянина, 1959). Особенно сильно уловы упали в 1972-1973 гг.
Общий вылов атлантическо-скандинавских сельдей неуклонно увеличивался с 1904 по 1930 г. (рис. 79), хотя, естественно, бывали кратковременные снижения. С 1930 по 1941 г. уловы, значительно колеблясь вокруг средней многолетней за этот период, держались примерно на одном уровне. Период второй мировой войны характеризуется некоторым снижением общего вылова. С 1947 г. вылов норвежской сельди опять стал быстро возрастать вплоть до 1955-1957 гг., когда он достиг максимума. В 1958 г. началось резкое падение уловов атлантическо-скандинавских сельдей, продолжающееся до настоящего времени (Marty a. Fedorow, 1963; Оstvedt, 1964). Некоторые исследователи (фstvedt, 1963, и др.), трактуют это снижение уловов только как результат следовавших друг за другом ряда неурожайных лет. Однако, как правильно отмечают Ю. Ю. Марти и С. С. Федоров, сводить причины падения промысла атлантическо-скандинавских сельдей только к появлению ряда неурожайных поколений, конечно, нельзя. Оно явилось результатом действия двух процессов: низкой величины пополнения, связанной с неурожайными поколениями последних лет, и интенсивной промысловой смертности. Последний фактор оказывает очень сильное воздействие как на половозрелую, так и на неполовозрелую части стада атлантическо-скандинавских сельдей.
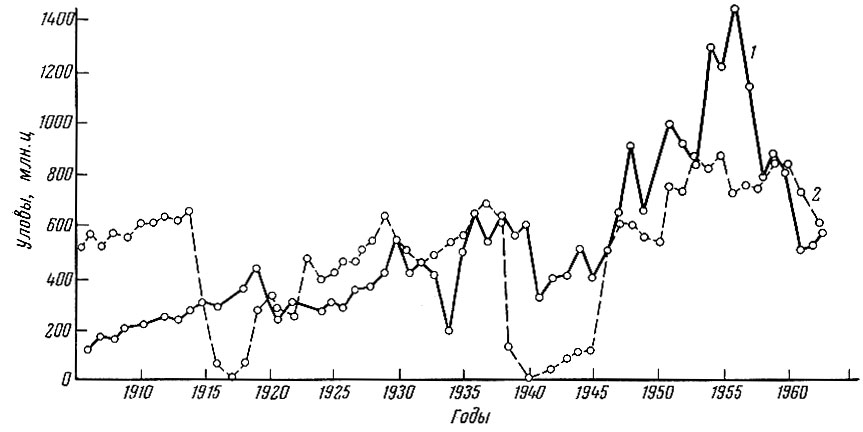
Рис. 79. Динамика уловов атлантическо-скандинавской сельди и сельди Северного моря (по данным Марти; Gushing a. Burd, 1957; Никольского и Беляниыой, 1959 и другим источникам): 1 - норвежские воды (атлантическо-скандинавская сельдь); 2 - Северное море и Ла-Манш (банковые сельди)
Уловы сельди Северного моря достигли максимума в 1913 г. - перед первой мировой войной, затем последовали падение уловов и новый подъем начиная с 1920 г. Период с 1920 по 1938 г. подобно тому, что мы наблюдали в случае с атлантическо-скандинавскими сельдями, может быть охарактеризован как относительно стабильный период с некоторыми колебаниями вокруг средней многолетней без тенденции к увеличению или уменьшению. С 1940 по 1945 г. в связи со второй мировой войной уловы сельдей Северного моря упали. По окончании войны они начали быстро возрастать, причем этот подъем продолжался до 1955 г., когда наметилось некоторое снижение. Однако если общая величина вылова сельди в южной части Северного моря возрастала до 1955 г., то вылов сельди Англией, промысел которой базируется главным образом на старших возрастных группах, начал снижаться уже с 1948 г. (Cushing a. Burd, 1957).
В настоящее время уловы сельдей Северного моря также находятся на низком уровне (Gulland, 1972).
Таким образом, величина вылова обоих исследуемых стад сельди в настоящее время, по-видимому, приблизилась к максимальной, которую при современной организации промысла (вылов значительного количества молоди и т. п.) уже не может компенсировать стадо рыбы и чтобы сохранить и восстановить его необходимо резкое снижение уловов. Естественно, что, когда в стадо вступает урожайное поколение, уловы могут увеличиваться, а при наличии неурожайных поколений уменьшаться.
У атлантическо-скандинавских сельдей отклонения максимальных и минимальных уловов от средних, допустимые для популяции, значительно больше, чем у североморской сельди. Это, видимо, связано с большей амплитудой урожайности отдельных поколений атлантическо-скандинавских сельдей.
Различные изменения наблюдаются и в структуре стада у анализируемых стад сельди. Анализ возрастного состава стада норвежской сельди за 50 лет показывает, что увеличение вылова с 150 тыс. в 1910 г. до 1500 тыс. т в 1954 г. не привело к изменению ни среднего, ни максимального возраста рыб в популяции (рис. 80). Закономерного омолаживания нерестовой популяции у норвежской сельди, несмотря на чрезвычайно возросшую интенсивность промысла, не происходит. Колебания среднего возраста, достигающие очень большой величины (рис. 81), видимо, в значительной степени определяются колебаниями урожайности отдельных лет. В известной степени на величине пополнения, несомненно, сказывается, как это правильно отмечает Марти (1956), и интенсивный вылов молоди, который еще сильнее снижает величину пополнения, особенно малоурожайных поколений.
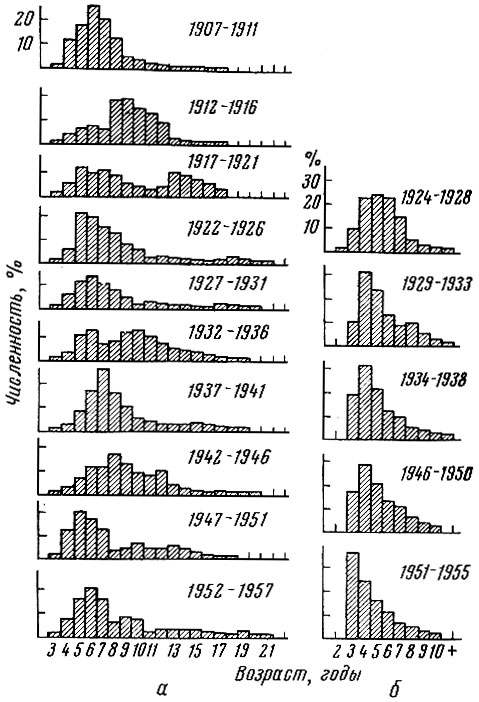
Рис. 80. Возрастной состав некоторых стад атлантической сельди (по пятилетиям) (по Никольскому и Беляниной, 1959; Marty, 1958; Gushing a. Burd, 1957): а - норвежская сельдь; б - сельдь Северного моря
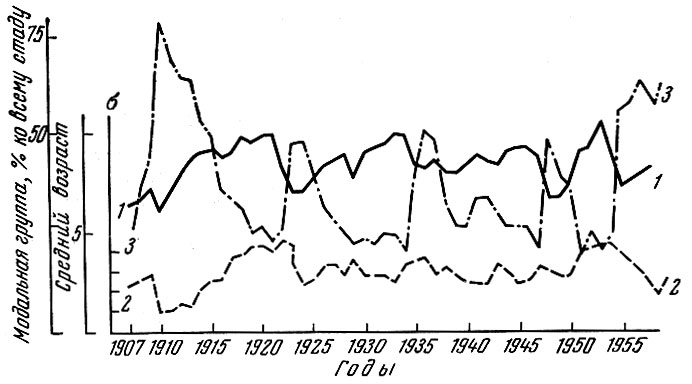
Рис. 81. Изменение некоторых показателей возрастного состава норвежской сельди (по Никольскому и Беляниной, 1959): 1 - средний возраст промысловых уловов; 2 - о возрастного состава стада; 3 - модальная возрастная группа (в %) по отношению ко всему стаду
Однако было бы преждевременным делать вывод, что увеличение вылова никак не отразилось на возрастном составе стада норвежской сельди. В 1955-1963 гг. в возрастном составе норвежской сельди имеется одна доминирующая возрастная группа (поколение 1950 г.), превышающая почти в 10 раз численность рыб любой другой возрастной группы. Несомненно, что эта особенность возрастной структуры стада связана в первую очередь с низкой урожайностью предыдущих и последующих поколений. Некоторое влияние на образование подобной возрастной структуры мог оказать промысел, как уже отмечалось, через уменьшение пополнения. Однако и в предыдущие периоды стадо норвежской сельди при значительно меньшей нагрузке промысла (в том числе и много меньшем вылове молоди) имело весьма сходную структуру возрастного состава.
Мы вычислили средний возрастной состав стада норвежской и южной североморской сельди по пятилетиям (см. рис. 80). Возрастной состав норвежской сельди в 1927-1931, 1947-1951 и 1952-1957 гг. имеет сходный характер.
Для более объективного суждения о характере изменений, происходящих в популяции, мы попытались вычислить некоторые показатели, ее характеризующие. В качестве показателей мы взяли средний возраст популяции по годам, процент модальной возрастной группы от всей популяции и среднее квадратическое отклонение возрастного состава нерестовой популяции. Ход кривой среднего возраста у норвежской сельди по годам и ход кривой а возрастного состава (см. рис. 81) оказывается сходным. При увеличении среднего возраста увеличивается и среднее квадратическое отклонение, и наоборот.
Попытаемся проанализировать полученные данные. Прежде всего отметим еще раз, что 10-кратное увеличение улова норвежской сельди с 1900 по 1958 г. не привело к закономерному изменению ни среднего возраста, ни амплитуды изменчивости возрастного состава популяции, ни процента модальной возрастной группы в стаде.
В последние годы у норвежской сельди при некотором повышении среднего возраста популяции резко уменьшается величина среднего квад-ратического отклонения, т. е. падает значение в популяции других возрастных групп, кроме модальной. Процентное отношение численности модальной группы ко всей популяции очень хорошо отражает относительную мощность урожайного поколения по отношению к другим, слагающим популяцию.
Отдельные пики на кривой процента модальной возрастной группы и их высота отражают вступление в промысловое стадо урожайного поколения, его мощность и - в известной степени - ход убыли. Среднее квадратическое отклонение отражает степень разброса возрастного ряда. Ход этого показателя у норвежской сельди может до известной степени свидетельствовать о наличии урожайного поколения в стаде, а в случае падения величины о при повышении среднего возраста, видимо, - о чрезмерном воздействии промысла, в первую очередь на младшие возрасты. В качестве предварительного, требующего проверки предположения можно указать, что одновременное повышение среднего возраста, снижение процента модальной группы и снижение а возрастного состава свидетельствуют о перенапряжении популяции сельди промыслом. Естественно, что это положение должно быть еще тщательно проверено.
Иная картина динамики возрастного состава стада наблюдается у сельдей южной части Северного моря.
Английские (Cushing a. Burd, 1957), бельгийские (Gillis, 1958) и французские (Ancellin, 1955) исследователи отмечают, что если предельный возраст сельдей южной части Северного моря по мере увеличения вылова не претерпевает существенных изменений, то средний возраст с 1941 по 1945 г. заметно возрос, а с 1948 по 1950 г. наблюдается прогрессивное омоложение популяции, после чего в течение 6 лет (1951-1956) средний возраст оставался стабильным.
Сравнение изменений среднего возрастного состава сельди Северного моря и норвежской сельди по пятилетиям (см. рис. 80) обнаруживает коренные различия. У норвежской сельди не удается отметить закономерного омоложения промысловой популяции, у сельди же южной части Северного моря очень четко выражена картина значительного омоложения возрастного состава.
Анализ динамики популяции сельди Северного моря показывает, что кривые среднего возраста популяции и среднего квадратического отклонения (рис. 82) также почти на всем своем протяжении имеют сходный характер. У сельди Северного моря колебания урожайности, видимо, много меньше, чем у норвежской сельди. Если у норвежской сельди относительная величина основной возрастной группы в популяции может варьировать в 60 раз, то у сельди южной части Северного моря только в 8-10 раз (Никольский, 1958).
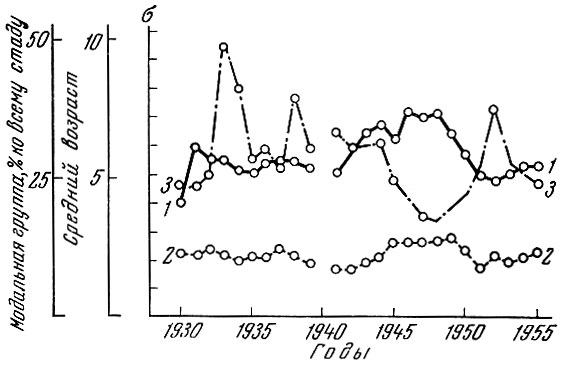
Рис. 82. Изменение некоторых показателей возрастного состава сельди юго-западной части Северного моря (по Никольскому и Беляниной, 1959): 1 - средний возраст промысловых уловов; 2 - о возрастного состава стад; 3 - модальная возрастная группа (в %) по отношению ко всему стаду
Существенные различия наблюдаются и в характере изменений, происходящих в темпе роста сравниваемых стад сельдей. У норвежской сельди не наблюдается закономерного ускорения или замедления роста (рис. 83). Повышение интенсивности рыболовства, видимо, не оказывает непосредственного влияния на изменение скорости роста норвежской сельди. Ускорения темпа роста, связанного с увеличением обеспеченности пищей в результате разрежения стада выловом, не наблюдается и регуляции пополнения через изменения роста, видимо, не происходит. Некоторые колебания темпа роста, по-видимому, определяются изменениями гидрологического режима, а тем самым продолжительностью периода нагула. Это согласуется и с тем, что более южные популяции норвежской сельди, живущие в условиях более длительного периода нагула, растут несколько быстрее, чем более северные популяции (Бирюков, 1956). В связи с тем что темп роста норвежской сельди не меняется в зависимости от разрежения стада, не меняется и возрастная структура пополнения, т. е. омоложения пополнения не происходит и темп пополнения сохраняется без значительных изменений (Sund, 1943).
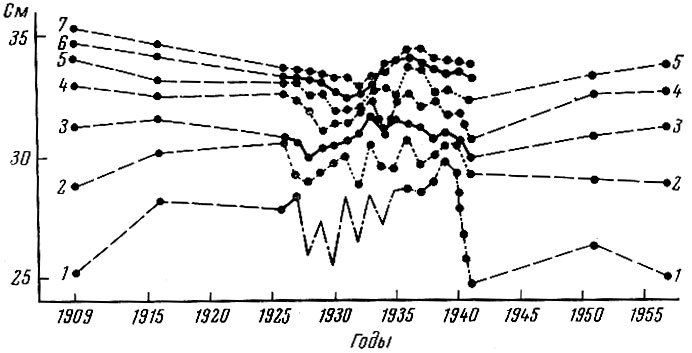
Рис. 83. Средняя длина одновозрастных особей норвежской сельди в разные годы (по Никольскому и Беляниной, 1959): 1-4-летки; 2-5-летки; 3-6-летки; 4-7-летки; 5-8-летки; 6-9-летки; 7-10-летки
Совсем иная картина наблюдается у сельди южной части Северного моря. У этой сельди по мере разрежения популяции темп роста значительно ускоряется (рис. 84) (Krefft, Schubert a. Sahrhage, 1955; Cushing a. Burd, 1957; Gilis, 1958; Ancellin, 1958). Неуклонное ускорение темпа роста у сельди южной части Северного моря наблюдалось с 1930 по 1954 г. С 1954 г. этот процесс у большинства возрастных групп приостановился. Возможно, что популяция сельди Северного моря перешла в последнее время в такое же состояние, что и норвежская сельдь, когда увеличение интенсивности вылова уже не приводит к увеличению обеспеченности пищей и не меняет структуры пополнения. Изменения гидрологического режима в сторону похолодания и тем самым сокращение периода нагула у сельди южной части Северного моря, по данным голландского исследователя Зийлстра (Zijlstra, 1958, 1963), могут в отдельные неблагоприятные годы вызвать замедление темпа роста так же, как это наблюдается и у норвежской сельди. Однако общая тенденция ускорения темпа роста в связи с разрежением популяции у сельди южной части Северного моря до 1955-1956 гг. была выражена очень четко и вызвала омоложение промыслового стада в период 1948-1950 гг. С 1955 по 1960 г. темп роста менялся слабо, а с 1960 г. наметилось некоторое ускорение, видимо, под влиянием климатических причин.
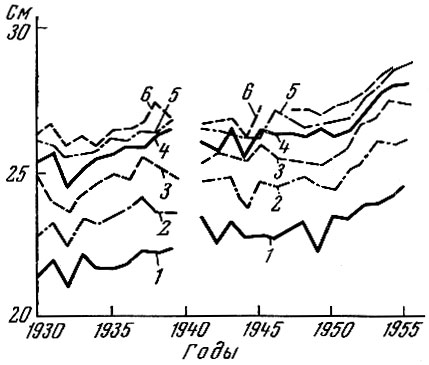
Рис. 84. Средняя длина одновозрастных особей сельдиюго-западной части Северного моря по годам (по Никольскому и Беляниной, 1959): 1 - 3-летки; 2 - 4-летки; 3 - 5-летки; 4 - 6-летки; 5 - 7-летки; 6 - 8-летки
Таким образом, если у норвежской сельди при разрежении популяции наступление половой зрелости не ускоряется, а следовательно, и не увеличивается темп пополнения, у сельди южной части Северного моря эти явления имеют место.
В данном случае изменение возрастного состава стада в сторону омоложения в результате интенсивного вылова - это в первую очередь результат омоложения возрастного состава пополнения, а не результат изъятия рыб старших возрастов. Это наглядно показывает сравнение динамики возрастного состава норвежской и южной североморской сельди.
Различаются эти стада также по изменениям плодовитости: у норвежской сельди плодовитость одноразмерных групп за последние годы заметно не изменилась, а у сельди южной части Северного моря она заметно возросла (Никольский и Белянина, 1959).
Весьма ценный материал о характере влияния вылова на структуру популяции промысловой рыбы дают анализ размерного и возрастного состава промысловых уловов древнего человека и сопоставление их с современными. В древних поселениях, где рыба была одним из основных объектов хозяйства, видимо, не происходил отбор в пищу только крупных экземпляров; лишь в поселениях, где рыба как пищевой продукт имела подчиненное значение (например, в южных сельскохозяйственных районах), как правило, в пищу поступали наиболее крупные особи (например, сазан и сомв городище близ станицы Елизаветинской на Кубани). Сравнивая размерный и возрастной состав рыб в промысловых уловах древности с современными, мы можем с известной долей точности выявить, какое влияние оказывает вылов на структуру популяции промысловой рыбы (Никольский, Соколов, Цепкий, 1972).
Как я уже писал (Никольский, 1935), размерный состав стерляди в уловах в Волге за последние два тысячелетия весьма сильно изменился. Размеры стерляди в современных уловах резко уменьшились и количество крупных рыб (в %) в промысловых уловах относительно сократилось.
Однако максимальные размеры добываемых рыб остались в настоящее время такими же, что и два тысячелетия назад (рис. 85).
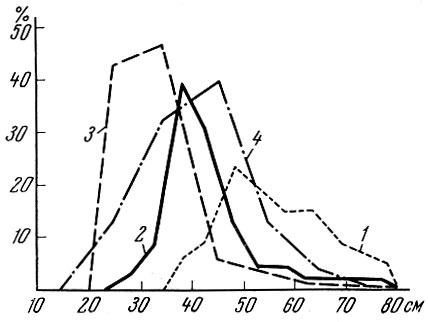
Рис. 85. Соотношение различных размеров стерляди в Одоевском городище (1), в среднем течении Волги (2), у устья Ветлуги (3) и в городище Больмеры (4) (по Никольскому, 1935, с добавлениями)
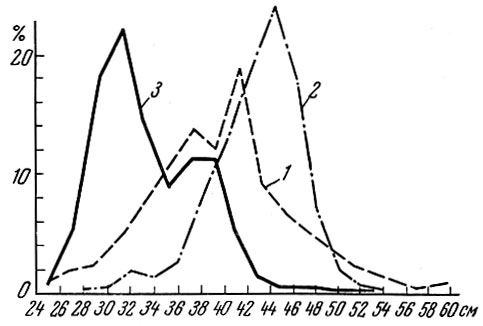
Рис. 86. Размеры леща (по Лебедеву, 1961): 1 - Древний Псков, I - XIII вв. н. э.; 2 - Псковское озеро, 1927 г.; 3 - Псковское озеро, 1951 г
Сравнивая размеры сазана и язя, добывавшихся в водоемах низовья Амударьи пять тысячелетий назад, с современными размерами этих рыб в дельте этой реки, мы видим, что средние и максимальные размеры этих рыб почти не изменились, причем это наблюдается при довольно значительной величине современного вылова этих рыб (табл. 92).
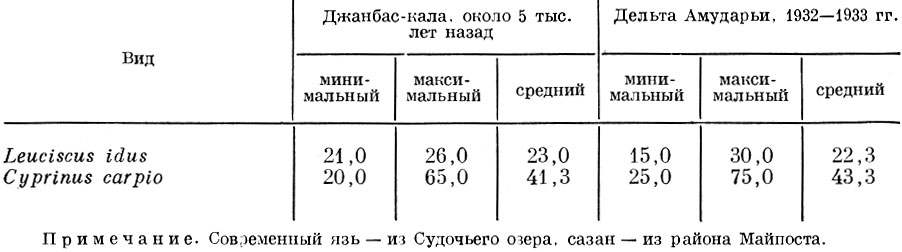
Таблица 92. Размерный состав сазана и язя в уловах амударьи, см
При сравнении размерного и возрастного состава леща из Псковского озера около 600 лет назад с современным (Лебедев, 1961) мы увидим резкое уменьшение средней длины современного леща (рис. 86). Одновременна с этим наблюдается и ускорение темпа роста. Однако в 1927 г., по данным Всесоюзного научно-исследовательского института озерного и речного рыбного хозяйства (ВНИОРХ), вылавливались значительно более крупные лещи даже по сравнению с ископаемыми, но, видимо, это связано с применением иных орудий лова. На темпе роста псковского леща в 1927 г. такое увеличение рыб старших возрастов в уловах не отразилось. Сходная картина наблюдается и с лещом Аральского моря. Средние размеры леща Аральского моря резко изменяются (правда, об ископаемых рыбах наши данные очень скудны) (табл. 93). В результате интенсификации промысла средние размеры леща в уловах снижаются и, наоборот, при уменьшении интенсивности рыболовства, как это было, например, в годы гражданской войны, средний размер леща в уловах быстро возрастает.

Таблица 93. Средняя длина леща (до конца чешуйного покрова) юга Аральского моря
Изменение размерного и возрастного состава леща Арала, видимо, почти не связано с изменением темпа роста. Этим аральский лещ отличается от псковского леща (рис. 87).
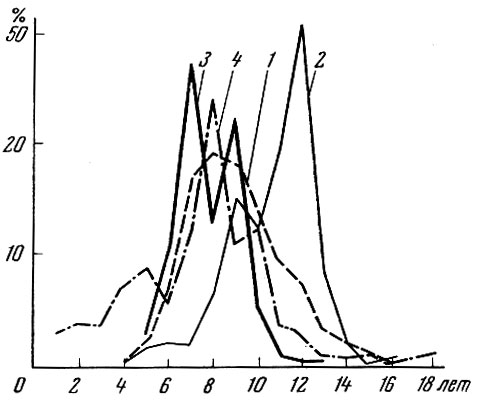
Рис. 87. Возрастной состав промыслового леща из различных водоемов: 1 - Древний Псков, X-XIII вв. н. э.; 2 - Псковское озеро, 1927 г.; 3 - Псковское озеро, 1951 г.; 4 - Сямозеро, 1946-1948 гг
Однако в связи с изменением режима Арала в последние годы темп роста аральского леща замедлился при продолжающемся снижении его уловов.
Сравнение размерного состава промысловых уловов амурского верхогляда Erythroculter erythropterus Bas. 2000 лет назад и в настоящее время с несомненностью показывает (табл. 94), что хотя в результате нерационального вылова и сокращения поголовья стада современные уловы верхогляда резко упали, но размерный состав в большинстве районов существенно не изменился.

Таблица 94. Размеры (в см) верхогляда в промысловых уловах
Число подобных примеров изменения или стабильности структуры популяций промысловых рыб по сравнению со структурой уловов человека древности можно было бы значительно увеличить. Большой материал по этому вопросу приведен в книге В. Д. Лебедева (1960).
Однако уже сделанный нами беглый анализ размерного и возрастного состава промысловых уловов древности и современных показывает, что в одних случаях и при относительно слабом воздействии промысла происходили весьма значительные изменения в структуре облавливаемой популяции, в других (например, у верхогляда) даже чрезмерная интенсивность промысла, связанная с резким снижением уловов, не привела к изменению размерной структуры стада.
"Не менее характерные изменения в структуре популяции в связи с интенсификацией рыболовства или его запуском подмечены в результате таких грандиозных экспериментов, как запуски рыболовства во время первой и второй мировых войн.
Как видно из изложенного, изменения величины вылова, часто очень значительные по масштабам, весьма по-разному отражаются на структуре популяции и других свойствах различных промысловых видов. У одних рыб структура популяции резко меняется, у других изменения оказываются практически незаметными.
Приведенные данные достаточно ясно показывают, что в отношении не только пресноводных, но и многих морских рыб рыболовство стало мощнейшим фактором, оказывающим на стадо промысловых рыб очень сильное влияние.
Изымая часть стада, вылов меняет условия жизни как того вида, который является объектом добычи, так и видов, находящихся во взаимосвязях с данным промысловым видом.
Уменьшение численности промыслового вида связано с уменьшением выедания им кормовых объектов и ослаблением противоречивых отношений с другими видами на почве питания сходной пищей. Освобождение кормовой базы часто приводит к усиленному размножению других видов, не являющихся объектами столь интенсивного промысла. Такая замена отмечена как в морских, так и в пресных водах. В качестве примера можно напомнить замещение более ценных видов камбал менее ценными на банках залива Петра Великого (Моисеев, 1946, 1946а), где в результате интенсивного промысла таких ценных видов, как, например, Limanda aspera (Pall.) , резко возросла численность менее ценных видов камбал; Hippoglossoides elassodon Jordan and Gilbert и др., а также таких сорных рыб, как морские лисички сем. Agonidae.
Сходная картина наблюдается и в пресных водоемах. Во многих озерах в результате чрезмерной интенсивности вылова леща его кормовая база часто используется ершом. В озерах бассейна Амура чрезмерный вылов сазана привел к резкому увеличению численности малой косатки Liocassis brashnikowi Berg.
Уменьшение численности промыслового вида отражается и на видах, для которых промысловый вид является жертвой. Разрежение популяции ухудшает условия охоты вида-хищника. Часто нарушение при облове структуры стаи облегчает хищнику охоту за своей добычей - мелкими стайными рыбами, добыча которых в стае для хищника затруднена. Так, при облове кошельковыми неводами стай анчоусов и сардины после того, как часть стаи выловлена, на оставшуюся часть стаи набрасываются держащиеся около нее тунцы и акулы и очень быстро ее уничтожают. В этом случае увеличение промысловой смертности приводит к увеличению смертности от хищника. Зачастую разреживание популяции связано с ускорением роста оставшихся особей, а тем самым с более быстрым выходом их из-под воздействия хищников, ибо, как известно, интенсивность воздействия хищников по мере роста жертвы резко падает: чем быстрее растет рыба, тем обычно скорее она выходит из-под воздействия хищников. Таким образом, вылов изменяет взаимосвязи организмов в экосистеме водоема, иногда приводя к ее коренной перестройке.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'