Селективность орудий и способов лова
Всякий вылов в той или иной степени и форме селективен. В некоторых случаях селективность рыболовства может быть выражена очень слабо, и практически ее можно не учитывать, в других (и таких случаев большинство) селективность рыболовства сильно отражается на структуре стада. Селективность рыболовства определяется двумя моментами: отбирающей способностью орудий рыболовства и поведением рыб (особей разного возраста, принадлежащих к одному и тому же виду и особей одного и того же возраста, но находящихся в разном биологическом состоянии).
Характер отбирающего действия орудия рыболовства есть результат взаимодействия орудия и рыбы. Селективное действие, как известно, зависит от размеров орудия, его конструкции, материала, из которого орудие сделано, места, времени, способа применения. Характер селективного действия определяется видовым и размерным составом облавливаемого стада, биологическим состоянием рыб в популяции (зрелость половых продуктов, жирность, упитанность, накормленность, характер поведения и др.).
При конструировании каждого орудия или разработке способа лова нужно учитывать свойства рыб, для добычи которых орудие или способ предназначены. Размер орудия, ячеи сетного полотна, из которого конструируется орудие, его материал и цвет должны выбираться из расчета на то, какую рыбу предполагается ловить (ее форма, подвижность, органы чувств). При выборе орудия лова должен учитываться и сезон лова. То же относится и к способам лова. Например, при лове на свет необходимо учитывать видовые особенности реакции рыбы на свет, характер органа зрения, в первую очередь спектр видимости, сезонную динамику освещенности и сезонную динамику реакции рыбы на свет, связь реакции рыбы на свет с ее биологическим состоянием.
Селекция орудиями и способами лова проявляется как в отборе рыб определенного вида или видов из облавливаемого скопления особей разных видов, так и в отборе определенных особей из одновидовой популяции. В пределах популяции одного вида формы селективности могут быть весьма разнообразными. Рассмотрим лишь основные из них:
отбор рыб определенного размера;
отбор рыб определенного пола и зрелости;
отбор рыб определенной упитанности и жирности;
отбор рыб разной накормленности.
Об отборе рыб определенного размера путем применения орудий лова с ячеей той или иной величины накоплена обширная литература.
Увеличение размеров ячеи кутца трала приводит к сдвиганию кривой размерного состава рыб в улове вправо (рис. 63). Уменьшение ячеи, наоборот, приводит к сдвигу размерного состава улова влево.
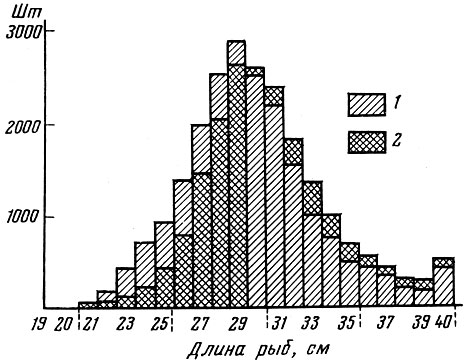
Рис. 63. Влияние размера ячей на длину вылавливаемых рыб (по Graham, 1939, 1948): 1 - ячея нормального размера; 2 - ячея большого размера
Из трех типов сетных орудий рыболовства (отцеживающие, ловушки и объячеивающие) размеры ячеи сетного полотна в орудиях первых двух типов определенным образом влияют на характер левого крыла кривой размерного состава облавливаемой популяции, пропуская мелких рыб. При изменении размеров ячеи сетного полотна изменяется минимальный размер залавливаемых орудием рыб. На правую часть размерного ряда облавливаемой популяции изменение размеров ячеи (естественно, до определенного предела) не оказывает прямого влияния, и обычно правая ветвь кривой размерного состава рыб в уловах отражает размерный состав более крупных рыб в облавливаемой популяции. При лове объячеивающими орудиями существует прямая связь между минимальными, средними и максимальными размерами добываемых рыб и размерами ячеи сетного полотна (речь идет об одностенных сетях).
На примере размерного состава балтийской салаки, добытой мелкоячейным тралом и жаберными сетями с ячеей 10 мм, отчетливо видно разное отбирающее действие отцеживающего и объячеивающего орудий лова (рис. 64). Накоплен очень большой материал по селективному действию различных орудий рыболовства. Особенно детальные исследования (в том числе совместно учеными разных стран) проведены по изучению селективности кутцов тралов. Показана селективность кутцов разной конструкции, с разным способом посадки полотна, кутцов, сделанных из разного материала, из безузловой дели и дели обычного типа. Весьма детально исследована селективная роль различных покрытий. Особенно много материала накоплено в изданиях Международного совета по изучению моря и Рыбохозяйственной комиссии Северо-Западной Атлантики. Подробный анализ селективности тралов дан А. И. Трещевым (1964).
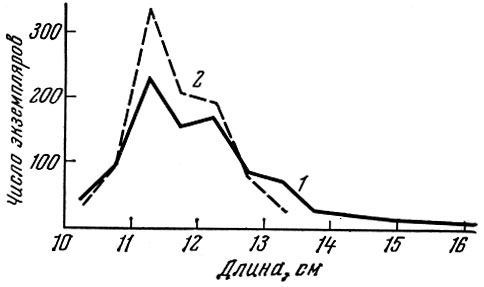
Рис. 64. Размерный состав балтийской салаки из ставных сетей и трала (по Никольскому и Беляниной, 1959): 1 - трал; 2 - килечная сеть
Связь размеров ячеи и длины рыбы, добываемой сетью, выражается простой формулой
(8)
где:
а - размер ячеи;
l - размер рыбы;
К - коэффициент, который зависит от вида и экстерьера рыбы и меняется с возрастом рыбы и изменением ее упитанности (Баранов, 1960).
Ф. И. Баранов (1936, I960) приводит специальный график, характеризующий связь длины рыбы с разной высотой тела и ячеи орудия лова (рис. 65).
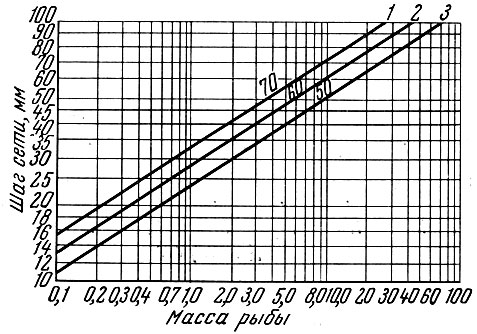
Рис. 65. Соотношение между массой рыбы, высотой тела и радиусом ячеи (по Баранову, 1933): 1 - окунь; 2 - судак; 3 - плотва
У большинства рыб, как это правильно отмечают Киплинг (Kipling, 1957) и Н. К. Пятерикин (1958), соотношение размеров улавливаемой сетью рыбы с размерами ячеи в объячеивающих орудиях в значительной степени зависит от формы, точнее "конусности" тела рыбы. Конусность Н. К. Пятерикин (1958) выражает следующей формулой:
| β = | C2 - C1 | |
| S |
(9)
где:
С2- больший обхват передней части тела перед спинным плавником;
С1- меньший обхват передней части тела за жаберными крышками;
S - расстояние между обхватами.
В пределах соответствия обхватов тела периметру ячеи сети рыба обычно и залавливается сетью. В свою очередь обхват (обычно наибольший) не остается постоянным в течение жизненного цикла рыбы и в течение года; он варьирует с изменением упитанности, степени наполненности кишечника пищей, зрелости половых продуктов. В связи с этим варьируют и размеры добываемых сетью рыб (Farran, 1936; Le Cren, 1951; Kipling, 1957 и др.).
Эта зависимость справедлива только в отношении рыб с относительно равномерным, пологим подъемом спины - таких, как сельди, голец Salvelinus sp., большинство сигов, плотва, язь, окунь, треска и многие др. У рыб с резким подъемом спины за затылком (например, карась, спаровые и др.) эта зависимость искажается.
Под коэффициентом селективности (Ks) (selection factor) обычно понимается отношение длины рыбы (Z), 50% которой проходит через ячею, а 50% остается в орудии лова, к размерам ячеи (В), т. е.
| KS = | l 50% | |
| B |
(10)
Существенное влияние на размерный состав улова оказывает и материал, из которого сделано орудие лова. Так, например, при лове бычка-кругляка Neogobius melanostomus (Pall.) в Азовском море драгами с ячеей в кутце 20 мм при кутце из капроновой нитки 34/18 ловятся более крупные бычки, чем при кутце с тем же размером ячеи, но из хлопчатобумажной нитки 20/12 (табл. 82).
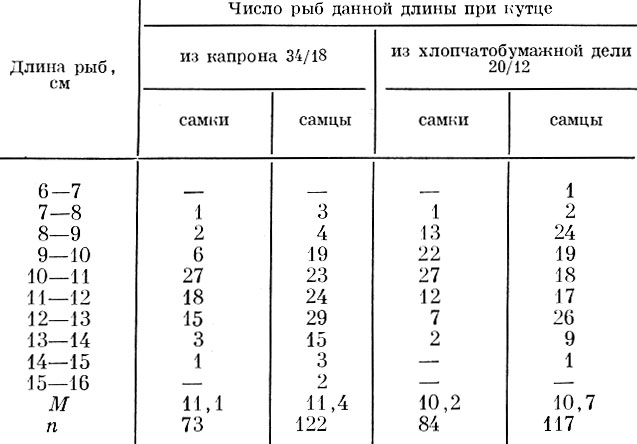
Таблица 82. Размеры добываемых бычков Neogobius Melanostomus (Pall.) Драгами с кутцами из разной нитки (ячея 20 мм) (по Ионасу, 1960)
Значительные различия в размерах нерки, добываемой жаберными сетями из искусственных и естественных волокон, отмечены Тоддом и Лар-киным для вод Британской Колумбии (Todd and Larkin, 1971).
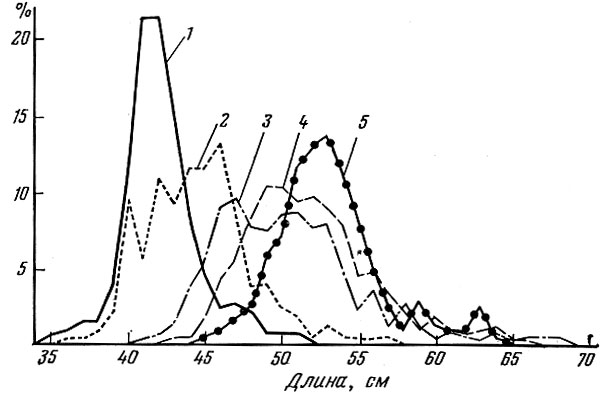
Рис. 66. Распределение судака Lucioperca lucioperca (L.) в % по размеру в уловах сетями с разной ячеей (ячея измерялась в растянутом виде - 2а) (по Havinga a. Deel-der, 1949): 1 - 86 мм; 2 - 92-96 мм; 3 - 100 мм; 4 - 105 мм; 5 - 110 мм
Размеры добываемой рыбы зависят также от толщины нитки в орудии лова.
Существеннейшее селективное значение, особенно в отцеживающих орудиях, имеет характер посадки сетного полотна. От того, как "раскрывается" при движении ячея отцеживающего орудия, зависит величина прилова молоди.
Сильно меняется селективное действие тралов и других отцеживающих орудий и в зависимости от величины улова. Чем больше улов, тем, как правило, шире варьирует фактор селективности, тем больше прилов маломерной рыбы. Это хорошо показал Боль (Воhl, 1964) в отношении камбал.
Из общей закономерности соотношения между размерами ячеи орудия лова и залавливаемой рыбы есть много исключений.
Некоторые виды рыб улавливаются в сеть не только путем объячеивания передней частью тела; часть особей зацепляется за сетное полотно челюстными костями (щуки, судаки, сиги и др.), зазубренными лучами плавников (сазаны, караси, многие сомы и т. д.) или жучками (осетр, стерлядь, лопатонос). В улове судака жаберными сетями в Нидерландах, по данным Havinga a. Deelder (1949), основную массу улова составляют рыбы, объячеивающиеся передней частью тела, некоторое количество обычно более крупных особей зацепляется за сеть челюстными костями (рис.66). Это же явление мы наблюдали при попадании щук в килечные сети в бассейне Амура и судаков и крупных окуней в мелкоячейные сети на Учинском водохранилище. Зацепляясь челюстными костями, в ставные сети попадают обычно только хищные рыбы в момент охоты за добычей. Особенно резко увеличивается процент пойманных таким способом рыб в период интенсивного нагула. Желудки у них обычно пустые или наполненные слабо. Мирные рыбы, например сиги и другие, редко цепляются за сеть таким образом. Однако при лове плавными сетями, когда сеть находится в движении, или при бота льном лове, когда рыбу загоняют в сеть, мирные рыбы тоже могут зацепляться за" сетное полотно в довольно значительном количестве. Иногда процент добытых подобным образом молодых особей "незаконной" длины бывает весьма значителен.
Так, по данным Л. И. Денисова (1951), прилов молоди судака в Цимлянском водохранилище рамовыми капроновыми сетями с ячеей 36 мм составлял 64,3% (по числу штук от всего улова судака), сетями с ячеей 40 мм - 40,0% и с ячеей 50 мм - 22%. По данным того же исследователя, в Цимлянском водохранилище в сети, выставляемые в русле Дона, попадает от 1 до 15 шт. мелкой стерляди, которая зацепляется за сеть главным образом спинными жучками.
Существенное отбирающее действие оказывает также размер крючков и наживки при лове рыбы на крючковую снасть, причем во многих случаях имеют значение не столько механические причины, сколько характер восприятий рыбой крючка и наживки (Thurow, 1964).
Значительно меньше исследован вопрос о селективности рыболовства, зависящего от поведения рыб разного размера. Как показано Ю. Е. Милановским и В. А. Рекубратским (1960) на примере обыкновенного гольяна, рыбы разного размера по-разному реагируют на один и тот же раздражитель. При появлении раздражителя сначала из кормового скопления уходят более крупные особи, затем средние и потом мелкие. Таким образом, орудие лова имеет больше возможностей захватывать более мелких рыб, чем крупных. Эта форма селекции по размерам еще плохо изучена, между тем при конструировании орудий рыболовства на нее необходимо обратить самое серьезное внимание.
Существенным источником отбора рыб определенного размера является их распределение (вертикальное или горизонтальное).
"Сезонно-территориальная" селективность - это такая форма селективности, когда состав улова отличается от состава стада в результате того, что рыбы разного размера держатся в разных участках водоема или слоях воды. Например, повышенный прилов молоди пикши на Канинской банке или молоди бихайбы Lutianus synagris (L.) в прибрежных водах Кубы объясняется не отбирающим действием орудий лова, а концентрацией здесь особей младших возрастов.
Как указывает М. Б. Шефер (1958), более крупные особи тунцов держатся глубже, чем мелкие, поэтому и улавливаются они обычными орудиями хуже и для их промысла требуются специальные орудия лова. То же самое относится к треске. Так как крупная треска меньше связана с дном, чем мелкая (это отмечал еще И. И. Месяцев, 1939), то тралом она улавливается хуже, чем мелкая. Донные тралы отбирают более мелкую рыбу.
Разница в составе дневных и ночных траловых уловов часто бывает связана с суточными вертикальными миграциями.
Такое же значение имеет разница в горизонтальном распределении крупной и мелкой рыбы. На знании мест концентрации молоди палтуса по Тихоокеанскому побережью Северной Америки основывается установление запретных для рыболовства мест.
В некоторых случаях, когда крупные и мелкие особи одного и того же вида питаются разной пищей, селекция по размерам может получаться при лове на разную наживку или крючками разного размера. Отбирающее действие оказывает рыболовство и на рыб разного пола. Наиболее обычная форма отбора рыб того или иного пола сетными орудиями рыболовства связана с разницей в размере самцов и самок. Как хорошо известно, сетной лов ходовой воблы в Северном Каспии дает больший процент самок, а сетной лов ходовой нерки в приустьевых районах р. Фрезер дает больший процент самцов (Peterson, 1954). При лове нерки в Бристольском заливе жаберными сетями ячеей 51/2 дюйма на нерестилища пропускается часто вдвое больше самок, чем самцов (Mathisen, 1962). Подобное явление отмечено у бычка-кругляка, при промысле которого крупноячейными драгами процент более мелких самок в уловах оказывается меньше самцов. Так, в уловах драг ячеей в кутце 20 мм самки составляют от 29 до 43%. Нормальное же соотношение полов у бычка-кругляка равно 1 : 1 (Ионас, 1960).
Существенное значение в отборе рыб того или иного пола имеет лов в местах, где концентрируются самцы или самки. Так, при промысле ходовых дальневосточных лососей в начале хода в уловах обычно значительно больший процент составляют самцы, а в середине хода самки. Регламентация лова в тот или иной период хода должна проводиться с очень большой осторожностью, дабы не нарушить необходимого соотношения полов у нерестующего стада.
Очень яркий пример селекции рыб одного пола у морских окуней (Sebastes) Северной Атлантики приводит В. П. Сорокин (Sorokin, 1963). Он показал, что интенсивный облов обыкновенного морского окуня Sebastes marinus L. приходится на момент, когда самки концентрируются в местах вымета личинок. Такой вылов преимущественно самок, по мнению В. П. Сорокина, уже сказался на эффективности размножения этого вида. Интенсивный облов окуня-клювача Sebastes mentella Travin, наоборот ведется в тех местах и в то время на местах нагула, где и когда концентрируются преимущественно самцы. Это также нарушает нормальное соотношение полов у этих, по-видимому, моногамных рыб.
По мнению Альма (Aim, 1959), более длительное пребывание самцов обыкновенного окуня на местах нереста, которые интенсивно облавливаются сетями, большая подвижность самцов по сравнению с самками приводят в некоторых озерах к повышенному вылову самцов этого вида.
Разная интенсивность вылова самцов и самок может явиться результатом неодинаковой реакции особей разных полов на один и тот же раздражитель. Так, самки каспийской кильки с текучими половыми продуктами не привлекаются на свет, вследствие чего в это время ускользают от облова (Шубников, 1959).
Практически, по-видимому, при промысле каждого вида рыбы один из полов вылавливается в относительно меньшем количестве, чем другой, но в большинстве случаев, вероятно, эта разница невелика и не отражается на воспроизводстве популяции. В некоторых случаях, когда промысел в большом количестве отбирает особей того или другого пола, несомненно, может нарушаться нормальный ход воспроизводства.
Селективное действие рыболовства может сказываться и в отборе рыб определенной стадии зрелости половых продуктов. В этом отношении, видимо, большее значение имеет лов объячеивающими орудиями. Поскольку сети отбирают рыб определенного экстерьера, то может получиться так, что из стада будут отбираться рыбы с наиболее отвисшим брюшком, т. е. с более развитыми половыми железами. Селективность объячеивающих орудий по этому признаку отмечает Ле Крен (Le Cren, 1951) в отношении обыкновенного окуня.
Очень существенное селективное действие на популяцию оказывает вылов, отбирая рыб определенной упитанности.
На примере сельди Северного моря Фарран (Farran, 1936) показал, что размеры рыбы, залавливаемой сетью с одной и той же ячеей, меняются в течение года, поскольку существует прямая зависимость между коэффициентом упитанности и обхватом рыбы как наибольшим, так и за жаберными крышками, а коэффициент упитанности весьма существенно изменяется в течение года. Так, например, в сети ячеей 12-19 мм попадает сельдь при коэффициенте упитанности 0,95 и длиной от 29,38 до 33,20 см, а при коэффициенте упитанности 1,05 в те же сети попадают рыбы длиной от 21,73 до 31,10 см. Как удалось показать, рыбы мельче среднего размера, улавливаемого сетью, оказываются более упитанными по сравнению с одноразмерными рыбами средней упитанности в популяции. Наоборот, особи, которые крупнее модальной группы, оказываются менее упитанными по сравнению со средней упитанностью одноразмерных рыб в популяции (Белянина, 1961; Никольский и Белянина, 1959).
Если размеры ячеи в сети подобраны для вылова модальной размерной группы особей (как в приводимом ниже примере с салакой), такая сеть берет рыб модальных размеров той же упитанности, что и упитанность рыб в популяции (рис. 67). Однако более мелкие рыбы (левая часть размерной кривой сетного улова) имеют более высокую упитанность, чем средняя упитанность рыб этих размеров в популяции, а рыбы правой части кривой размерного ряда в сетном улове имеют меньшую упитанность, чем средняя упитанность рыб этого размера в популяции.
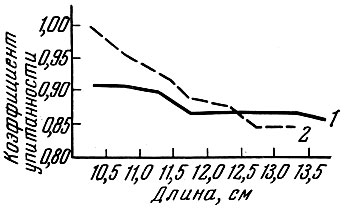
Рис. 67. Упитанность салаки Clupea harengus membras (L.) (по Фультону) из трала (1) и ставных сетей (2) (по Никольскому и Беляниной, 1959)
Если размеры ячеи в сетях ниже оптимальных для модальной группы рыб в популяции, то сеть с такой ячеей ловит особей с упитанностью ниже средней упитанности одноразмерных рыб в популяции (рис. 68). Если размеры ячеи в сетях выше оптимальных для модальной размерной группы в популяции, то такая сеть отбирает особей с более высокой упитанностью, чем средняя упитанность одноразмерных рыб в популяции.
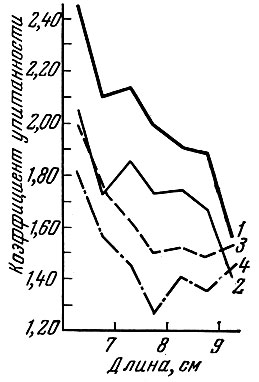
Рис. 68. Упитанность ерша ЛсеПпа cernua (L.) из невода и ставных сетей (по Никольскому и Беляниной, 1959): 1 - из неводных уловов (по Фультону); 2 - уловов (по Фультону); 4 - из неводных уловов (по Кларк); 3 - из сетных из сетных уловов (по Кларк)
Подобные наблюдения проведены нами на ряде морских и пресноводных рыб. Сравнение коэффициентов упитанности (по Фультону и Кларк) особей салаки одной и той же длины, добытых тралом и жаберными сетями с ячеей 10 мм, позволило установить следующую закономерность (см. рис. 68): рыбы, составляющие в сетном улове модальную группу (11,5 - 12,5 см), по упитанности практически почти не отличаются от рыб этих размеров, добываемых тралом. Сравнение упитанности добытых сетями более мелких рыб с одноразмерными особями из трала показывает, что сети отбирают более упитанных мелких рыб из популяции. Обратная картина наблюдается в отношении более крупных рыб: упитанность особей из сетей несколько ниже, чем у одноразмерных особей из трала. Таким образом, сеть отбирает более упитанных мелких рыб и менее упитанных крупных. Так как у салаки, по наблюдениям Л. Е. Анохиной (1960), более упитанные особи плодовитее и жирнее, то оказывается, что из младших возрастов сеть отбирает более ценных особей, а из более крупных рыб - менее ценных.
Близкая картина наблюдается и у норвежской сельди: мелкие особи в уловах дрифтерными сетями оказываются упитаннее рыб средних размеров, а более крупные рыбы упитаны ниже, чем рыбы средних размеров. Сравнение упитанности одноразмерных особей норвежской сельди, добытых дрифтерными сетями с ячеей 24 и 32 мм (рис. 69), показывает, что рыбы, добытые сетями с более крупной ячеей, гораздо упитаннее, чем сетями с более мелкой ячеей. У рыб, добытых сетями с ячеей 24 мм, закономерность, отмеченная выше в отношении салаки и заметная у норвежской сельди, добытой сетями с ячеей 32 мм, не выражена. По-видимому, установленная закономерность проявляется тогда, когда размер ячеи близок к оптимальному для модальной группы рыб в популяции. При лове же сетями с более мелкой ячеей отбора может и не происходить: у наиболее крупных размерных групп сельди, добытых сетями с ячеей как 24, так и 32 мм, коэффициент вариации упитанности оказался наибольшим.
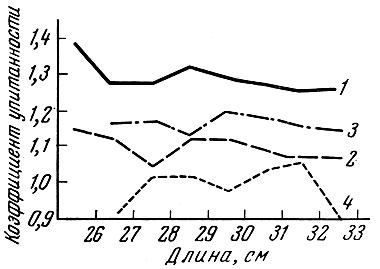
Рис. 69. Упитанность атлантической сельди из дрифтерных сетей с разным размером ячеи (по Никольскому и Беляниной, 1959): 1 - сельди из сетей с ячеей 32 мм (упитанность по Фультону); 2 - то же (упитанность по Кларк); 3 - сельди из сетей с ячеей 24 мм (упитанность по Фультону); 4 - то же (упитанность по Кларк)
Это же наблюдается и в отношении отбирающего действия объячеивающих орудий при лове карповых рыб: у востробрюшки Hemiculter leucisculus sbsp., добытой в оз. Далай-Нор (табл.83), так же как и у салаки, более мелкие особи оказываются упитаннее одноразмерных особей, добытых неводом, а более крупные рыбы из сетных уловов менее упитанны, чем одноразмерные особи из неводных уловов. Сходную картину мы обнаружили у амурской востробрюшки Hemiculter leucisculus. Добытые килечными сетями мелкие особи значительно упитаннее более крупных. У особей же, добытых отцеживающими орудиями лова, коэффициент упитанности по мере роста увеличивается, что, видимо, характерно для естественных популяций этого вида (Никольский, 1956а).
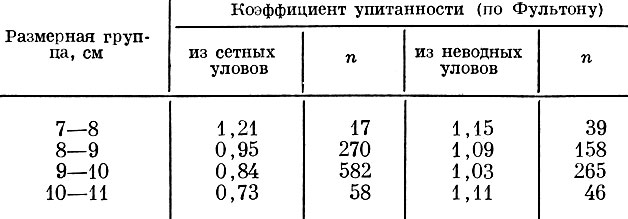
Таблица 83. Упитанность вострое рюшки из неводных и сетных уловов
У амурского чебака Leuciscus waleckii Dyb. и уссурийского пескаря Gnathopogon chankaenensis Berg, добытых килечными сетями, более мелкие особи также упитаннее более крупных (табл. 84) в отличие от того, что наблюдается в естественных популяциях (Никольский, 1956а).

Таблица 84. Упитанность амурского чебака и уссурийского пескаря, добытых килечными сетями с ячеей 10 мм
Анализ упитанности уклеи Alburnus alburnus (L.), добытой в Учинском водохранилище ставными сетями с ячеей 10 и 18 мм и закидным неводом, показывает, что неводом ловится более упитанная уклея, чем сетями с ячеей 10 мм. Так, у рыб длиной 9-10 см, добытых неводом, коэффициент упитанности по Фультону равнялся 1,37 и по Кларк 1,29, а у рыб того же размера из сетей с ячеей 10 мм коэффициент упитанности по Фультону был 1,15 и по Кларк 0,96. Уклея, добытая сетями с ячеей 18 мм, оказывается более упитанной, чем в неводе и в сетях с ячеей 10 мм. Упитанность уклеи длиной 10-12 см из сетей ячеей 18 мм была по Фультону 1,64 и по Кларк 1,43, а из сетей с ячеей 10 мм соответственно 1,06 и 0,98. Упитанность уклеи в сетях с ячеей 18 мм превышает также упитанность уклеи в неводных уловах, т. е. сети с ячеей 18 мм отбирают из популяции более упитанных рыб (табл. 85).
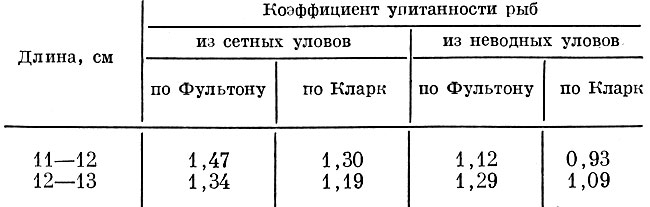
Таблица 85. Упитанность уклеи, добытой в учинском водохранилище сетями с ячеей 18 мм и неводом
Сравнение упитанности плотвы Rutilus rutilus (L.), добытой в Учинском водохранилище сетью с ячеей 18 мм и неводом, показывает,что сеть отбирает из популяции менее упитанных рыб (табл. 86).
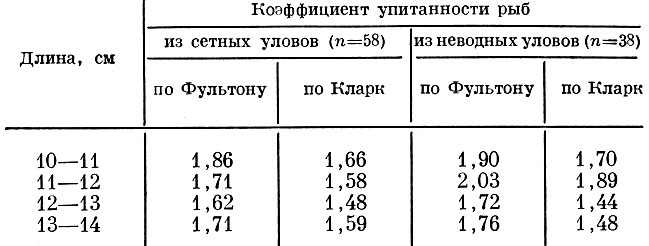
Таблица 86. Упитанность плотвы, добытой в учинском водохранилище ставной сетью с ячеей 18 мм и неводом
Таким образом, сеть с ячеей 18 мм оказывается "мала" для облавливаемой популяции плотвы; отбирая более мелких рыб, она изымает менее упитанных. Следовательно, закономерности селективного действия жаберных сетей, выявленные при анализе материала по сельдевым рыбам, подтвердились и на карповых.
Сопоставление упитанности окуня Perca fluviatilis L., добытого ставными сетями с ячеей 18 мм и неводом, показывает (табл. 87), что сеть с ячеей 18 мм отбирает из популяции значительно более упитанных рыб.

Таблица 87. Упитанность окуня, добытого в учинском водохранилище ставной сетью с ячеей 18 мм и неводом
У ерша Acerina сеrnuа (L.) упитанность рыб, добытых сетями с ячеей 10 мм, много ниже, чем рыб, добытых неводом (см. рис. 68). Это указывает на то, что сеть с ячеей 10 мм "мала" для учинской популяции ерша. Как в неводных, так и в сетных уловах более крупные особи ерша характеризуются более низким коэффициентом упитанности.
Рассмотрим селективное действие рыболовства на рыб разной накормленности.
Лов крючковыми снастями в период нагула безусловно отбирает из нагульного скопления рыб определенной накормленности.
Перед окончанием периода нагула, когда более упитанные и жирные рыбы прекращают питаться, а более худые интенсивно кормятся, лов наживными орудиями отбирает из популяции менее упитанных и жирных рыб. Селекция рыб разной накормленности осуществляется не только при лове на наживку.
В объячеивающие орудия челюстными костями попадают только голодные хищные рыбы. Сытые рыбы таким образом улавливаются крайне редко. Сытые рыбы обычно осторожнее голодных и легче избегают орудий лова. Поскольку голодные рыбы часто ведут себя иначе, чем сытые, то, естественно, что они могут иначе облавливаться и отцеживающими орудиями, и ловушками. Так, например, как показано Б. П. Мантейфелем и Л. А. Чаяновой (1960), кошельковый лов сельди у Северной Исландии ведется во время питания ее красным калянусом Calanus finmarchicus, причем в кошельковые невода попадаются рыбы только с наполненным пищей кишечником. Рыбы с пустыми кишечниками в кошельковые невода не попадаются.
Я счел необходимым остановиться на рассмотрении форм селективного действия рыболовства потому, что, как мне представляется, и экономическое, и биологическое значение этого вопроса очень велико, а закономерности селективного действия рыболовства на популяции промысловых рыб мы знаем еще очень плохо.
Совершенно недостаточно изучены генетические аспекты селективности рыболовства, а между тем отбор из стада упитанных и более быстрорастущих рыб, несомненно, может оказывать отрицательное влияние на воспроизводительные свойства и продуктивность популяций промысловых рыб (Никольский, 1966).
Пока рыболовство не было интенсивным, его отбирающее действие можно было игнорировать. При современной же и все возрастающей интенсивности рыболовства влияние его селективного действия на качественный состав облавливаемой популяции нельзя недоучитывать. Как я попытаюсь показать дальше, путем селективного воздействия рыболовства человек может управлять структурой популяции рыб, а тем самым и ее продуктивностью.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'