Закономерности изменений структуры популяции под воздействием вылова
Как мы видели выше, вылов так же, как и изменения интенсивности естественной смертности, часто связан с определенными изменениями в структуре популяции промыслового вида. Изменяются возрастной состав, рост, возраст полового созревания и предельный возраст особей, соотношение полов и ряд других свойств популяции.
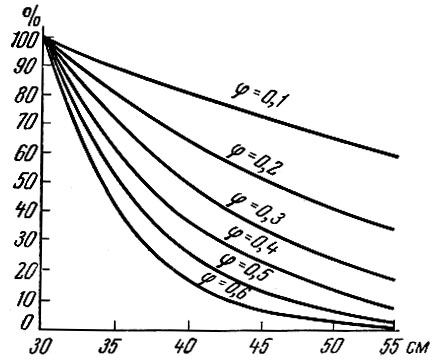
Рис. 88. Кривые населения при различных коэффициентах естественной убыли (φ) (по Баранову, 1918)
Теоретически селективным выловом можно изменить структуру популяции в очень значительной степени, даже довести ее до такого состояния, что популяция потеряет способность самовоспроизводиться.
Выше были рассмотрены закономерности, которым подчиняется изменение структуры популяции в результате изменения соотношения пополнения и остатка и возрастной структуры того и другого. Было показано, что при изменении интенсивности смертности, если она носит неселективный характер, т. е. погибают особи в том же соотношении, в каком они находятся в популяции, и сохраняется стабильным соотношение величины пополнения и остатка, - структура популяции не изменяется. Структура популяции при неселективной смертности изменяется, если меняется соотношение пополнения и остатка или темп роста особей, т. е. структура пополнения и предельный возраст рыб в стаде.
И в настоящее время широко распространено мнение, что возрастная структура стада промысловой рыбы является непосредственной функцией интенсивности рыболовства (в том числе неселективного) и что по изменениям кривой возрастного состава облавливаемого стада можно непосредственно судить об интенсивности рыболовства. Впервые эту точку зрения развил подробно Ф. И. Баранов (1918,1925, 1962). Ф. И. Баранов принимает, что:
1) "кривая населения", т. е. кривая численности рыб разного возраста в стаде, идентична кривой смертности;
2) кривая улова отражает кривую распределения рыб по размерам и по возрасту в водоеме. Правая часть кривой улова подобна кривой населения, а левая есть результат отбора применяемой ячеей определенной величины;
3) число ежегодно погибающих рыб промыслового размера равняется числу ежегодно подрастающих рыб минимальной длины независимо от вида кривой населения;
4) для каждой популяции существует своя оптимальная допустимая величина вылова. Поэтому основной метод рзгулирования рыболовства - это установление определенной нормы вылова;
5) интенсивность лова влияет на форму кривой населения и коэффициент К (коэффициент убыли). Она отражается на распределении в улове рыб разного возраста (длины и веса);
6) кривая, отвечающая различной интенсивности убыли (различный коэффициент годичной убыли φ), различна (рис. 88). Чем выше интенсивность промысла, тем круче кривая населения. При увеличении интенсивности промысла (от φ = 0,1 до φ = 0,6) соответствующим образом меняется кривая населения - кривая смертности.
В основе этих представлений Ф. И. Баранова лежит принцип сложных процентов*. В своих теоретических представлениях Баранов (1918, 1925) исходит из положения, что молоди в водоеме всегда достаточно и что пополнение стада определяется кормовой базой, которая освобождается в результате вылова определенной части взрослого стада.
* (Мы здесь не останавливаемся на математическом выражении представлений Ф. И. Баранова и других ученых. Этот вопрос подробно рассмотрен во многих работах. Можно сослаться хотя бы на Вестенберга (Westenberg, 1955).)
Томпсон (Thompson, 1937), исходя из представлений Баранова, построил арифметическую схему изменения структуры стада под влиянием рыболовства с интенсивностью в 50 и 80 %. Эту схему без серьезных критических замечаний повторили Э. С. Рессел (1939), И. Вестенберг (1957) и В. С. Ивлев (1959, 1961).
Схема Томпсона показана в табл. 95.

Таблица 95. Схема Томпсона
На втором примере Томпсон показывает, что, по его мнению, произойдет, если интенсивность промысла изменится с 50 до 80%. Как и в первом случае, на модельном примере (табл. 96) создается при первом взгляде впечатление, что происходит сокращение возрастного ряда вследствие якобы выпадения старших возрастных групп.
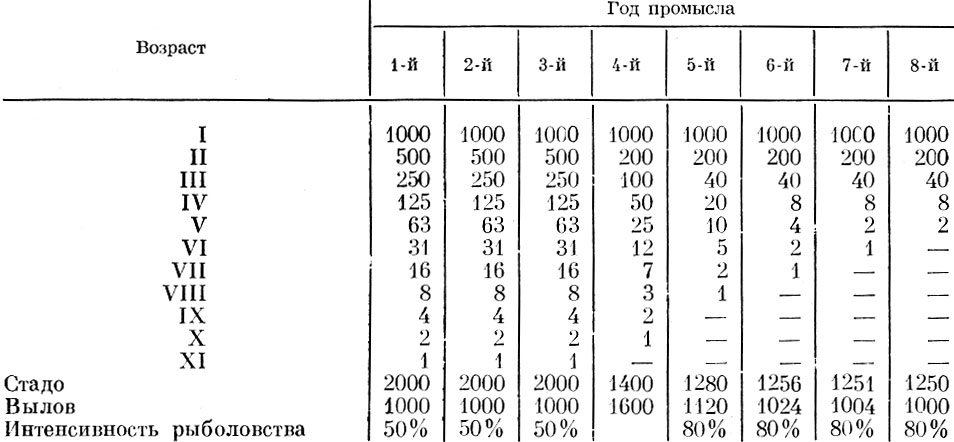
Таблица 96. Изменение структуры стада в случае изменения интенсивности рыболовства с 50 до 80% (по Томпсону)
В основу расчетов Томпсона так же, как и Баранова, положен принцип сложных процентов. Принимается, что из пополнения в N рыб изыг мается промыслом р рыб. К концу сезона промысла остается (1-p)N. Рыбы, родившиеся на год раньше, были обловлены дважды. Из рыб, сохранившихся от облова в прошлом году, на второй год также будет выловлена часть р. Число оставшихся рыб после этого вторичного облова будет равняться (1-p)2N. На третий год облова эта величина будет равна (1-р)3N и т. д. Численность каждого годового класса будет уменьшаться по убывающей геометрической прогрессии с знаменателем (1-р). При этом величина всего стада к началу рыболовного сезона будет:
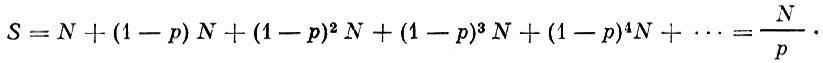
Таким образом получается, что численность стада обратно пропорциональна интенсивности рыболовства. Чтобы не возвращаться еще раз к рассмотрению приведенных Томпсоном модельных примеров, отметим, что, как я уже указывал (Никольский, 1961а), сокращение возрастного ряда в расчетах Томпсона получается искусственно вследствие "округления" цифр. В примере Томпсона, приведенном в табл. 95, вылов рыб в возрасте 2-4 лет составляет 80, а рыб 5 лет - 100%. В приведенном примере до старших возрастов доживали бы доли рыб, но при доведении исходной численности до десяти тысяч - миллиона или более соответственно удлинился бы и ряд возрастного состава стада. Так, например, 80% от двух будет 1,6, а принимается 2, т. е. округляется, причем в сторону завышения, и ликвидируется то незначительное абсолютное количество особей старших возрастов, которое на самом деле будет сохраняться. Омоложение популяции в примере, приводимом Томпсоном и повторенном Вестенбергом и Ивлевым, достигается не в результате интенсификации промысла, а вследствие непропорционально большого пополнения, которое принимается одинаковым (1000 рыб) в случае изъятия и 50, и 80%. В табл. 95 и 96 Томпсон принимает вылов рыб старших возрастов равным не 80, а 100%, т. е. получается уже селективный облов с повышенным выловом рыб старших возрастов.
Вестенберг (1955) правильно отмечает, что Томпсон при определении оптимальной интенсивности рыболовства в приводимых примерах не учитывает ни естественной смертности, ни индивидуального роста рыб.
Приводимые Томпсоном примеры отражают частный случай, который может быть в искусственном водоеме.
Остановимся теперь на рассмотрении основных положений теории рыболовства, принимаемых Барановым, Грэмом, Томпсоном и другими исследователями.
I. Является ли кривая населения одновременно и кривой смертности? Ф. И. Баранов и его последователи отвечают на этот вопрос утвердительно, а Г. Н. Монастырский (1949) - отрицательно. Как же обстоит дело в действительности?
Кривая населения - это как бы равнодействующая сложного процесса рождения, роста и убыли. Любая часть кривой отражает все эти процессы. Даже если мы возьмем все стадо (см. выше), а не только половозрелую часть популяции, то колебания урожайности часто (у рыб как с флюктуациями, так и без флюктуации) будут приводить к тому, что предыдущий годовой класс (например, 7-летки) будет многочисленнее последующего (например, 6-леток). При этом, если следовать Баранову, величина смертности получится с обратным знаком. Так как в природе не существует постоянной величины пополнения (никак не связанной с остатком, как это принимает Ф. И. Баранов), то приводимый им пример можно рассматривать как абстракцию, которая обычно в природе не существует и может иметь место в отдельных редких случаях. Конечно, при разреживании стада оно в какой-то мере компенсирует уменьшение численности путем повышения интенсивности воспроизводства, но это не значит, что нулевая точка кривой населения на оси ординат остается постоянной.
Смертность является одной из составляющих, которые определяют кривую населения, но она определяет кривую населения во взаимодействии с другими процессами - роста и урожайности поколений, а не изолированно.
Как говорилось выше, величину смертности можно определить, анализируя судьбу отдельных поколений, сопоставляя их численность из года в год в общих уловах или в уловах на единицу рыболовного усилия. Судить же по возрастному составу стада об интенсивности смертности можно было бы лишь в том случае, если бы, как это принимается в примерах Ф. И. Баранова (1918) и Томпсона (1937, 1950), величина пополнения была из года в год постоянной. Если у рыб флюктуации выражены слабо, то по среднему многолетнему возрастному составу нерестового стада, зная средний многолетний возрастной ряд пополнения, можно приблизительно определить осредненную величину общей смертности. Нужно, однако, при этом иметь в виду, что предельный возраст рыбы меняется в зависимости от возраста наступления половой зрелости, и вылов, разреживая стадо и ускоряя рост и наступление половой зрелости, иногда может через изменение возраста половой зрелости менять и максимальный возраст, достигаемый рыбой. В малочисленных поколениях продолжительность жизни особей бывает короче, чем в поколениях многочисленных.
Таким образом, отвечая на поставленный вопрос, я должен сказать, что кривая населения не аналогична кривой смертности. Смертность входит одним из элементов, которые определяют кривую населения.
II. Подобна ли правая часть кривой улова (в отцеживающих орудиях рыболовства) кривой населения? Этот вопрос нельзя решать абстрактно. Конечно, всякое орудие рыболовства в какой-то степени селективно. Как показал Томпсон (1917), даже при лове сельди кошельковым неводом может иметь место в некоторых случаях известная селективность (более крупные рыбы в большем проценте избегают орудия). То, что рыбы старших возрастов осторожнее молодых и раньше реагируют на приближение опасности, также, несомненно, имеет определенное селективное значение. Однако при облове нерестовой популяции отцеживающими орудиями или ловушками возрастной, размерный и половой состав изымаемого улова часто бывает сходен с составом облавливаемой нерестовой популяции, т. е. популяции рыб, достигших половой зрелости. Примеры подобного рыболовства общеизвестны (кошельковый лов нерестовой сельди, лов ходовых лососей ставными и закидными неводами, лов закидными неводами многих карповых при нерестовой миграции и т. д.), причем при облове нерестовой популяции часто оказываются сходными с действительным возрастным и размерным составом нерестового стада не только правая, но и левая части кривой улова, но это именно кривая населения, а не кривая смертности. С другой стороны, при облове нагульных скоплений, например при облове трески тралом на кормовых площадях, обычно получается недобор рыб старших возрастов, которые, будучи осторожнее молодых и держась несколько выше над грунтом, легче избегают попадания в орудия лова.
Распределение рыб разного возраста и пола (см. выше) естественно может также оказывать влияние на состав улова и соответствие его структуры структуре облавливаемого стада. Поэтому нам представляется, что поставленный вопрос должен решаться применительно к каждому конкретному случаю, но в принципе структура улова отцеживающим орудием или ловушкой может быть сходной со структурой облавливаемого стада.
III. Соответствует ли число ежегодно подрастающих рыб числу ежегодно погибающих. Известно, что численность поколений в пределах одного и того же стада у видов с хорошо выраженными флюктуациями может варьировать в десятки раз. Даже у рыб с относительно малыми колебаниями урожайности, как камбалы, численность отдельных поколений может разниться в 6-10 раз. Конечно, во многих случаях численность стад промысловых рыб может в течение более или менее длительного времени удерживаться на более или менее стабильном уровне, но эта относительная стабильность достигается при помощи различных регуляторных приспособлений, позволяющих популяции приспособительно отвечать на изменение обеспеченности пищей. Улучшение обеспеченности пищей может сильно изменить соотношение пополнения и убыли.
IV. Имеется ли для каждой популяции своя оптимальная норма вылова? Каждый вид приспособлен к определенной, конечно, изменяющейся в тех или иных пределах величине убыли. В отношении видов, лучше защищенных от воздействия хищников, допустима меньшая величина изъятия, чем в отношении рыб, которые приспособились к большей смертности и у которых убыль стада легче компенсируется интенсификацией воспроизводства. Естественно, что норма вылова или контингент вылова даже для одного вида или популяции не постоянная величина. Она варьирует в зависимости от состояния популяции. Кроме того, необходимо подчеркнуть, что, как это отмечали Н. Л. Чугунов (1928), Грэм (Graham, 1948) и ряд других исследователей, норма вылова меняется в зависимости от того, какого размера и в каком возрасте вылавливается рыба и в какое время года осуществляется вылов (Бервальд, 1956). Ниже на этих вопросах я остановлюсь подробнее.
V и VI. Влияет ли сама по себе интенсивность вылова (при неселективном облове) на форму кривой населения? Выше мы показали, что изменение возрастной структуры нерестовой популяции это не результат прямого изъятия промыслом или гибели в результате других причин. Кривая населения меняется в результате ряда причин, а именно изменения соотношения величины пополнения и остатка и изменения структуры пополнения и остатка. Сама по себе меняющаяся интенсивность вылова при неизменном соотношении пополнения и остатка не влияет на структуру кривой населения.
Перейдем к рассмотрению исходных теоретических представлений, положенных Ф. И. Барановым и его последователями в основу своих построений.
И Ф. И. Баранов, и ученые, развивающие его точку зрения, исходят из представления, что основной момент, которым ограничивается численность пополнения стада, это недостаток корма, происходящий от того, что корма выедаются другими особями того же вида. По мнению Баранова, число выметанных икринок, как правило, во много раз больше числа мальков, которое может выжить. Причина гибели личинок заключается в том, что другие особи съедают их корма. Таким образом, в основе построений Баранова лежит представление о биотическом потенциале и сопротивлении среды, причем основным элементом среды, ограничивающим численность пополнения, являются особи того же вида.
Выше в главе об обеспеченности пищей этот вопрос рассмотрен подробнее и показана неправильность этой точки зрения.
Принятие Ф. И. Барановым за основу своих взглядов представлений о наличии постоянной перенаселенности, а также вытекающих из этих представлений понятий о биотическом потенциале и сопротивлении среды и определило весь дальнейший ход его построений. На этих теоретических положениях основываются и принятые Ф. И. Барановым построения и графики (см., в частности, рис. 88) постоянной величины пополнения вне зависимости от величины остатка. Эту же ошибку допустил и Томпсон в приведенных выше модельных арифметических примерах. Если следовать Баранову, то при облове рыбы с нерестовой популяцией третьего типа интенсификация вылова в любом случае должна в конечном итоге привести к перестройке типа нерестовой популяции из третьего во второй тип. В природе же подобная картина обычно не наблюдается.
Переход популяции из состояния, когда остаток превышает пополнение, в состояние, когда пополнение превышает остаток, в природе совершается довольно часто и вне зависимости от интенсивности вылова. Перестройка нерестовой популяции из одного типа в другой не обязательно является результатом изменения интенсивности вылова. Структура популяции, как было показано выше, при вылове любой интенсивности может оставаться неизменной, если не меняется соотношение величины пополнения и остатка.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'