Низкая обеспеченность пищей как причина смертности
Формы влияния обеспеченности пищей на выживание популяции весьма разнообразны. Обеспеченность пищей влияет на интенсивность смертности главным образом косвенным путем. Как говорилось выше, более быстрый рост связан с более ранним выходом из-под интенсивного воздействия хищников; хорошая подготовленность к зимовке (высокая упитанность и жирность) ослабляет воздействие паразитов и снижает смертность (например, карпа в прудах). Лучшая обеспеченность родительского стада пищей приводит к продуцированию им более жизнестойкого потомства. Наконец, в некоторых случаях ухудшение обеспеченности пищей приводит к растягиванию сезона нагула и ставит иногда популяцию в неблагоприятные температурные и другие абиотические условия (например, в неблагоприятные по кормности годы задержка хамсы в Азовском море на местах нагула приводит иногда к массовой гибели). Недостаток пищи может быть непосредственной причиной гибели рыб, как правило, только на ранних этапах развития. Так, при отсутствии подходящей пищи личинки салаки Clupea harengus membras L. погибают на четвертые - пятые сутки (Лисивненко, 1960), личинка сельди гибнет после нескольких безрезультатных бросков за пищей (Soleim, 1942).
Отмечается (табл. 79) также прямая связь мощности поколения трески и богатства летнего планктона в годы рождения поколения (Corlett, 1958).

Таблица 79. Биомасса летнего планктона в районе медвежьего острова и урожайность поколения трески (по Corlett, 1958)
Состояние и выживание личинок в зависимости от обеспеченности пищей в разные периоды нерестового сезона наглядно показано Шелбурн (Shelbourne, 1957) на примере личинок морской камбалы. При низкой обеспеченности пищей на этапе смешанного питания в уловах преобладают очень худые личинки и совсем нет упитанных; наоборот, при богатом развитии планктона количество упитанных, более жизнестойких личинок сильно возрастает (рис. 57).
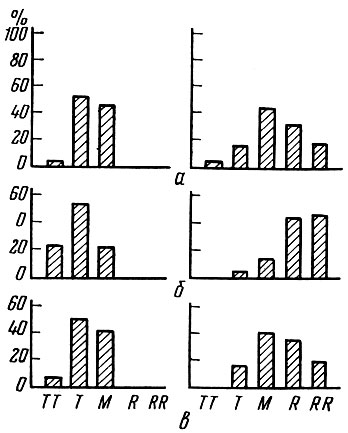
Рис. 57. Упитанность (в %) переходных стадий личинок камбалы при скудных и обильных скоплениях планктона (по Shelbourne, 1957): а - личинки перед полным рассасыванием желточного мешка, стадия 1 (d); б - начало постжелточной стадии 2 (а); в - общая переходная стадия 1 (d) + 2 (а). Слева - январские скопления (пищи мало); справа - мартовские скопления (пищи много). ТТ - очень худые; Т - худые; М - средние; R - упитанные; RR - очень упитанные
Однако прямолинейной связи между обилием кормовых организмов и выживанием личинок рыб нет. На графике, заимствованном из работы Р. М. Павловской (1963), мы провели наряду с прямой, намеченной автором, действительный ход зависимости величины поколения хамсы от количества корма (рис. 58); в пределах 4-8 тыс. экз. зоопланктеров на 1 м2 корреляции между численностью кормовых объектов и величиной поколения не наблюдается; при численности зоопланктеров от 8 до 12 тыс. экз. существует почти прямая корреляция между величиной кормовой базы и численностью поколения; наконец, при численности зоопланктеров от 12 до 16 тыс. экз. корреляции опять почти не наблюдается. Таким образом, имеются определенные как минимальные, так и максимальные пороговые концентрации, при которых численность поколения уже не зависит от обилия корма.
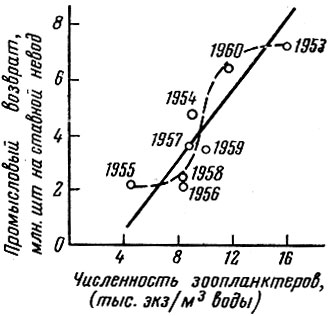
Рис. 58. Величина промыслового возврата поколений и численность кормовых для личинок хамсы организмов в июле (по Павловской, 1963). Пунктирная линия проведена нами
Очень четко выражена связь между численностью личинок салаки и количеством кормовых организмов (Лисивненко, 1961). В годы с богатым развитием планктона численность личинок салаки оказывается более высокой (рис. 59).
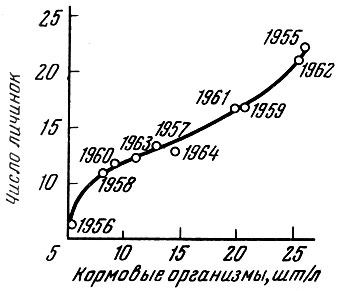
Рис. 59. Зависимость между количеством кормовых организмов в 1000 см3 воды и числом личинок салаки на 10 мин лова (по Лисивненко, 1961)
У черноморской хамсы также отмечена четкая связь условий нагула личинок на первых этапах внешнего питания и урожайности поколения (Павловская, 1963) (см. рис. 58).
С другой стороны, для ряда черноморских рыб, принадлежащих к средиземноморскому фаунистическому комплексу, обеспеченность пищей на ранних этапах онтогенеза достаточно высокая и сильно не отражается на их выживании (Дехник, Дука и Синюкова, 1970).
Очень большое значение в выживании личинок на первых этапах активного питания имеет состояние личинок и в первую очередь запас желтка у них (его количество и качество). Это отмечено у многих рыб. Например, у днепровского осетра Acipenser guldenstadti Brandt из более крупных икринок выводится более крупная и более жизнестойкая молодь. В условиях эксперимента разница между выживанием личинок, полученных из более крупной и из более мелкой икры, составила 10,6% в пользу первых (Семенов, 1963).
Блекстер и Хемпел (Blaxter a. Hempel, 1963) показали, что личинки разных форм атлантической сельди Clupea harengus L., вышедшие из икринок большей массы и имеющие больший запас желтка, могут дольше переносить голодание и голодные личинки позднее теряют способность захватывать корм (рис. 60). Эти же авторы отмечают, что потомство от впервые нерестующих рыб не так долго переносит голодание, как потомство от повторно нерестующих рыб того же стада.
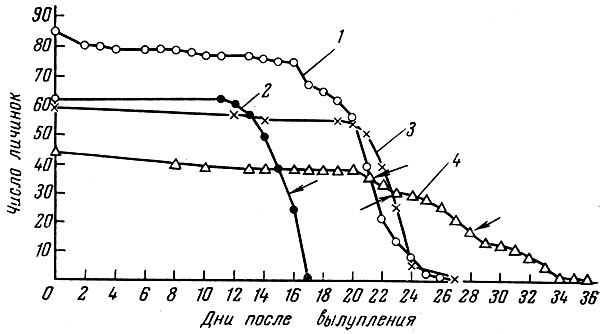
Рис. 60. Смертность личинок сельди в обычных экспериментах по голоданию из разных районов (по Blaxter a. Hempel, 1963): 1 - Доггер-банка; 2 - Киль; 3 - норвежская сельдь; 4 - район Дауне. Стрелками показана 50%-ная смертность
Разная выживаемость при голодании свойственна и разным локальным формам сельди. Обычно более северные формы приспособлены к перенесению более длительного голодания в личиночном состоянии, чем южные. Напомним, что северные формы обычно характеризуются меньшей относительной плодовитостью.
Намечается следующая закономерность: на краю ареала вида и фаунистического комплекса в целом абиотические факторы имеют большее-значение как причина смертности. Однако все факторы среды действуют во взаимной связи, и абиотические связи определяют величину смертности часто через изменения биотических связей.
По данным Г. И. Фроленко (1959), у леща гибель личинок, находившихся без пищи на этапе резорбции желтка (С1 по Васнецову), наступила на 6-е сутки после начала активного питания. Впервые сутки погибло 32,3% подопытных личинок. Последние голодавшие личинки погибли на 11-е сутки. На более поздних этапах гибель от голода начиналась позднее. Так, личинки, лишенные корма на этапе С2, начали гибнуть на 8-е сутки, личинки на этапе D1 погибли после 9-суточного голодания и на этапе Е прожили более 14 суток.
Отмечено, что при голодании на этапе смешанного питания у личинок медленнее рассасывается желточный мешок. Сходную картину В. С. Ивлев (1955) отметил у молоди воблы: более молодые личинки выдерживают голодание менее длительный срок, чем более старые; в среднем 10-дневные рыбки прожили без пищи в 3 раза меньший срок, чем 98-дневные. Однако отсутствие непосредственной гибели в результате голодания еще не говорит о выживании голодавшего поколения.
Как показали опыты Г. И. Фроленко (1959) с личинками леща на этапе C1 личинки, голодавшие одни и двое суток, в развитии почти не отставали от нормально питавшихся. При более длительном голодании рост замедлялся в соответствии с периодом голодания. Личинки леща, голодавшие 8 суток, не смогли возобновить питание и погибли.
Как показал еще Йорт (Hjort, 1914) и подтвердили очень многие исследователи, у ряда рыб (сельдь, треска и др.) величина поколения определяется на ранних личиночных этапах, особенно на этапе смешанного питания в период перехода на внешний корм. Условия жизни на этапе смешанного питания фактически решают судьбу поколения у многих видов рыб.
Нельзя согласиться с Марром (Магг, 1956) и Т. В. Дехник (1961), что высказанные Йортом мысли и сегодня являются только гипотезой. Они подкреплены многими новыми фактами (Гербильский, 1957; Гри-гораш, 1961; Грудинин, 1961; Дислер, 1953; Дементьева, 1958; Заика и Островская, 1973; Красюкова, 1958; Крыжановский, 1956; Костомарова, 1961; Лисивненко, 1960; Павловская, 1955, 1963; Панов, 1957 и др.).
Чем лучше обеспечена рыбка на этом этапе развития как желтком, так и внешней пищей, тем выше выживаемость. Рыбы с большим запасом кормовых ресурсов в желтке, естественно, при одной и той же обеспеченности внешней пищей оказываются в лучших условиях развития и выживают в большем количестве. Наличие большого запаса пищевых ресурсов в желтке на этапе смешанного питания позволяет лучше переносить недостаток пищи. Как установлено В. А. Григораш, на этапе смешанного питания у особей плотвы с большим запасом желтка существует суточная ритмика в питании внешним кормом, а особи с малым запасом желтка вынуждены потреблять пищу круглосуточно, что, естественно, ставит их в худшие условия выживания и делает более доступными хищникам.
Несомненно, что качество родителей в той или иной степени по-разному в разных условиях влияет на смертность личинок. Это хорошо показано В. И. Владимировым, К. И. Семеновым и В. Н. Жукинским (1965) на примере леща и тарани низовьев Днепра. В. В. Кормилин и В. П. Митрофанов (1972) установили, что у личинок балхашской маринки Schizothorax argentatus Kessl., перешедших на внешнее питание, смертность при голодании выше, если они получены от менее упитанных самок, обладающих большей интенсивностью обмена.
Однако нельзя согласиться с В. И. Владимировым (1964), который считает, что у ряда рыб "высокая смертность в период конца рассасывания желтка и начала питания экзогенной пищей объясняется не голоданием, а другими внутренними причинами". Эти "внутренние причины" В. И. Владимиров видит в том, что полноценные производители в нормальных условиях жизни дают тот или иной процент дефектного потомства, которое и погибает на определенных этапах развития. То, что у рыб в определенных условиях жизни может возрастать количество уродов в потомстве, - это факт, но то, что это может быть закономерным явлением, определяющим численность поколения в естественных условиях, - не соответствует фактам. Развитие - это процесс, протекающий в единстве внешнего и внутреннего. Интенсивность выживания определяется соответствием условий жизни и состояния организма, а не только какими-то внутренними причинами, переданными родителями потомству.
Поскольку выживание рыбы на этапе смешанного питания зависит и, видимо, в значительной степени от обеспеченности желточным питанием (т. е. кормами, полученными от родителей), то возможно, что по количеству и качеству желтка можно в какой-то степени составить представление о жизнестойкости потомства. Хорошим показателем пищевых качеств желтка может служить количество в нем жира и общего азота. Как известно, относительное количество общего азота в желтке икры у разных видов рыб изменяется сравнительно слабо. Так, по данным Нидгема (Needham, 1931), количество белка (в % сырой массы) колеблется у 15 видов рыб, о которых приводятся сведения, от 22,37 до 32,25%, тогда как количество жира - от 1,4 до 22,34%.
У шести видов рыб Черного моря, исследованных С. С. Виноградской (1954), количество азота (на сухую массу) колебалось от 6,33 до 14,31%, а жира - от 3,88 до 32,6%. Таким образом, по данным и Нидгема, и Виноградской, содержание белка (или общего азота) колеблется в 1,5-2 раза, а относительное количество жира в икре в 10 раз и более, т. е. жир является гораздо более изменчивым показателем, характеризующим состояние питательных веществ в желтке. И у одного и того же вида изменчивость содержания жира в икре на одной и той же стадии развития значительно больше, чем изменчивость общего азота.
Несомненно, что выживание разных видов рыб и разных поколений одного и того же вида на ранних этапах развития в значительной степени связано с обеспеченностью желточным питанием - как количеством, так и качеством желтка. Последнее же в значительной степени связано с условиями нагула и возрастом родителей.
Мне представляется, что неправа Т. В. Дехник (1961) в своей трактовке данных Сетте (Sette, 1943) и Алстрома (Alstrom, 1954) о роли обеспеченности пищей в выживании атлантической скумбрии и тихоокеанской сардины на этапе смешанного питания. Приводимые Сетте и Алстромом примеры мне кажутся достаточно убедительными. Примеры возрастания смертности на этапе смешанного питания можно дополнить данными о дунайской сельди (Владимиров, 1953), балтийской салаке (Лисивненко, 1960) и многими другими.
Марр (Маrr, 1956) выделяет среди морских рыб три типа "кривых выживания" (рис. 61). Первый тип характеризуется резким падением численности в течение короткого отрезка времени. У этих рыб, как говорит Марр, есть "критический период"*, когда смертность сильно возрастает. После этого короткого периода интенсивность смертности стабилизируется. Для характеристики подобного рода рыб Марр приводит данные Алстрома (Alstrom, 1954) о тихоокеанской сардине Sardinops sagax coerulea Girard, у которой массовая смертность приходится на 5-дневный возраст при
* (Термин "критический период" ("La periode critique"), насколько мне известно, впервые введен Фабр-Демергом и Биетри (Fabre-Demergue et Bietrix, 1897) для обозначения этапа смешанного питания у морских рыб. К своим выводам они пришли на основе аквариальных наблюдений за выращиваемыми личинками длине личинок около 3,25 мм. К этой же группе можно отнести и скумбрию.)
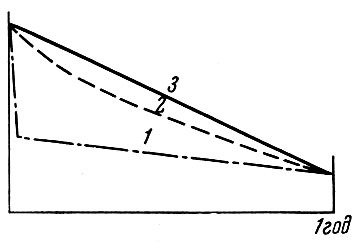
Рис. 61. Гипотетические кривые выживания от икры до годовика (по Маrr, 1956). Показатели выживания: 1 - очень низкие в течение критического периода; 2 - повышающиеся с постоянной скоростью; 3 - постоянные. По оси ординат отложено количество выметанных икринок; по оси абсцисс показано время от вымета икры до возраста 1 год
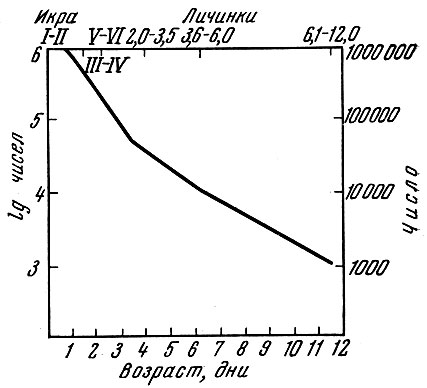
Рис. 62. Выживание икры и личинок черноморской хамсы (по Дехник, 1961). Римскими цифрами обозначены стадии развития личинок; арабскими длина в мм
Второй тип "кривой выживания" - это постепенное плавное уменьшение коэффициента смертности. Возможно, что такой тип кривой на ранних стадиях развития наблюдается у морской камбалы. Примером подобного типа кривой может служить кривая смертности (рис. 62) на ранних стадиях развития черноморской хамсы Engraulis encrassicholus ponticus Pus., приводимая Т. В. Дехник (1961). Наконец, третий тип, по Марру,- это постоянный коэффициент смертности на первом году жизни. Подобный тип кривой смертности, по его мнению, наблюдается у ряда рыб. В дальнейшем удастся установить, конечно, еще несколько "типов" кривых смертности, а вернее всего, что кривые смертности окажутся специфичными для разных видов и в то же время будут варьировать из года в год. Возможно, что в годы образования урожайных поколений кривая из первого типа с наличием "критического периода" будет превращаться в кривую второго типа, т. е. без "критического периода".
Несколько слов необходимо сказать о так называемых критических периодах. Как правило, на ранних этапах онтогенеза организм более стенобионтен, на более поздних - более эврибионтен. Часто наблюдается, например, что на одних этапах организм оказывается чувствительнее, например, к сотрясениям, на других - к колебаниям температуры. Так, икра дальневосточных лососей очень чувствительна к сотрясениям на стадии замыкания бластопора. В это же время она выдерживает и малые давления. Это и понятно, так как в это время икра развивается в грунте в спокойном состоянии. Усиление прочности оболочек и потеря острой восприимчивости к встряхиваниям возникают у икры примерно в тот период, когда в реке начинаются подвижки льда, и лед часто ложится на грунт при падении уровня воды. Вместе с тем на первых этапах после оплодотворения икры лососи сравнительно легко переносят значительные колебания температуры, так как в природных условиях температура в это время сильно колеблется.
К этапу формирования пигмента глаз амплитуда оптимальных температур несколько уменьшается, что связано с характером естественных изменений температуры в природной обстановке. Таким образом, когда рыбоводы говорят о "критических периодах" в развитии рыбы, они имеют в виду узкую приспособленность к определенному градиенту конкретного элемента или конкретных элементов среды, вырабатывающуюся как приспособительная реакция на относительную стабильность данного элемента среды в природной обстановке в период прохождения данного этапа развития.
Трактовать "критические периоды" как реализацию каких-то нарушений, полученных по наследству от родителей (Владимиров, 1964), неправильно. "Критические периоды"- это одна из форм проявления единства организма и среды, а не автогенетический процесс, проходящий вне зависимости от среды. Более подробно разбор представлений о критических периодах дан Е. Ф. Еремеевой (1967).
Известно, что у одних видов рыб колебания урожайности по годам выражены резко, у других слабо. По-видимому, у видов с резко выраженными флгоктуациями обычно выживание идет по кривой первого типа с наличием "критического периода", т. е. резкого перелома кривой. При этом максимальная смертность приходится обычно на один и тот же этап развития - на этап перехода с желточного питания на внешнее. У рыб со слабовыраженными флюктуациями на ранних этапах развития смертность тоже может резко увеличиваться, но коэффициент смертности обычно изменяется плавно. Если же и бывают резкие изменения коэффициента смертности, то они у одного и того же вида в разных водоемах и в разные годы обычно приходятся на разные этапы развития.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'