Влияние абиотических условий на смертность рыб
Роль абиотических условий в динамике численности и биомассы популяции как прямая, так и косвенная может быть весьма велика и разнообразна.
Косвенное воздействие сказывается через изменение кормовой базы, условий нагула, условий воздействия хищников и паразитов и т. д.
Прямое летальное воздействие абиотических условий, естественно, особенно сильно сказывается на ранних стадиях развития: на икре, свободных эмбрионах и личинках, которые совсем неподвижны или малоподвижны и при изменении условий жизни в неблагоприятную сторону не могут переместиться и погибают.
Летальное действие абиотических факторов на численность стада рыбы обычно более сильно сказывается на краю ареала вида или в случае резкого изменения условий, в частности в результате деятельности человека.
Процент икры, погибающей от неблагоприятных условий, у разных видов рыб весьма различен и часто очень сильно меняется по годам. Так, у красной оз. Дальнего, по данным Камчатского отделения ТИНРО, смертность икры на нерестилищах (по Крогиус, 1951) менялась следующим образом.
| Год | Погибло % от отложенной |
|---|---|
| 1938 | 99,6 |
| 1939 | 76,7 |
| 1940 | 82,7 |
| 1941 | 41,2 |
| 1942 | 28,5 |
| 1943 | 21,7 |
| 1944 | 21,0 |
| 1945 | 35,0 |
| 1946 | 51,0 |
Основной причиной смертности икры красной, видимо, является нарушение газового режима в буграх.
Для выживания пелагической икры морских рыб, развивающейся в поверхностных слоях воды, большое значение имеют волнения. Как установлено в отношении хамсы (Дехник, 1961; Павловская, 1963), волнения более четырех баллов вызывают массовую гибель икры. Особенно чувствительна икра к волнениям на этапе дробления. После шторма в Черном море от 30 до 50% икры хамсы обычно погибает.
По данным Т. В. Дехник (1961), смертность эмбрионов хамсы в Евпаторийском районе Черного моря в 1957 г. была 66,2, а в Прибосфорском районе в 1958 г. - 49,6%. Непосредственные причины смертности не установлены.
Смертность икры аральского леща на нерестилищах разного типа весьма различна (табл. 77). Она вызывается в первую очередь ночными кислородными заморами на нерестилищах типа наплавов. Икра, отложенная на урути Myriophyllum spicatum, где кислородный режим благоприятен, погибает в незначительном количестве.
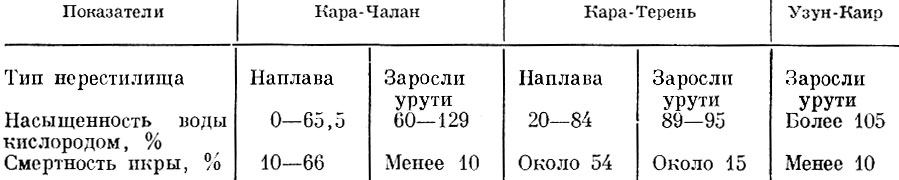
Таблица 77. Смертность икры леща на нерестилищах разного типа в северной части аральского моря в 1948 г. (по Бервальду, 1950)
У аральского сазана основная причина гибели икры на аральских нерестилищах - также кислородные заморы, возникающие главным образом в зоне наплавов и жесткой растительности и близ затопленных купа-ков тростника. На подобного типа нерестилищах насыщенность кислородом колеблется от 0 до 70, а смертность икры от 40 до 100%. На нерестилищах других типов, где кислородный режим благоприятен (60-110% насыщения 02), смертность икры сазана колебалась в 1948 г. от 6 до 20%.
Развитие в неблагоприятных для вида условиях приводит не только к гибели икры и личинок, но и к развитию уродливых особей. Так, по наблюдениям П. М. Коновалова (1950), при изменении солености воды отход икры при инкубации и количество уродливых личинок изменялись следующим образом (табл. 78).

Таблица 78. Отход икры аральской воблы rutilus rutilus aralensis berg. и процент уродливых личинок при инкубации при разных соленостях воды (по Коновалову, 1950)
Как видно, оптимальной для развития икры аральской воблы является соленость около 4-5‰. В воде выше этой солености отход икры и количество уродливых личинок возрастают. Однако известно, что икра аральской воблы развивается и при солености воды 11,6‰.
Таким образом, на стадии икринки и свободного эмбриона абиотические факторы могут вызвать массовую гибель икры и личинок. Это бывает в тех случаях, когда абиотические условия выходят за пределы границ, к которым вид оказывается на данном этапе развития приспособленным. В большинстве случаев плотность засева икры на растительность и закладки в гнезда не отражается на интенсивности гибели. Связи гибели икры аральского леща и сазана с плотностью засева ни нам в 1935 г., ни Э. А. Бервальду (1950) наблюдать не приходилось, однако это, конечно, не означает отсутствия связи между плотностью засева икры и ее выживанием у всех видов и во всех случаях. Примеры, приводимые И. Г. Фридлянд (1951) о сахалино-хоккайдской сельди, говорят об обратном. Блекстер и Холлидей (1963), ссылаясь на данные Э. Леа и Рунстрема (Runnstrom, 1941), отмечают, что если икра сельди у морского побережья Норвегии лежит слоем от 0,5 до 2,5 см, то смертность икры составляет от 43 до 80%. Однако Парриш с соавторами (Parrisch a. oth., 1959) даже в плотных скоплениях икры сельди у берегов Англии не наблюдали увеличения смертности. Блекстер и Холлидей (1963) на основании аквариальных наблюдений указывают, что у сельди есть оптимальные концентрации икры, при которых наблюдается наибольшее выживание. Как разреживание, так и увеличение густоты засева икрой усиливают смертность.
В некоторых случаях, видимо, встречающихся в природе довольно редко, массовая гибель икры может происходить в результате переуплотнения. Подробнее на этом вопросе мы остановимся ниже.
Гибель под воздействием абиотических условий резко возрастает под влиянием деятельности человека, в первую очередь в результате изменения режима рек гидросооружениями и загрязнения вод. Колебания уровня часто приводят к гибели всей отложенной икры, оказывающейся или на суше, или, наоборот, на больших глубинах при неблагоприятном температурном и гидрохимическом режиме. Даже суточные колебания уровня, когда икра оказывается на суше на короткий срок, вызывают ее повышенную гибель и появление большого количества уродливых личинок. Это отмечено для леща, сазана и ряда других рыб (Владимиров, Сухойван и Бугай, 1963; Владимиров, 1965). Гибель икры от обсыхания резко возрастает как в созданных на реках водохранилищах, так и в нижних бьефах плотин. Несоответствие режима колебаний уровня в результате деятельности гидросооружения оптимальному режиму в период нереста и развития икры рыб является в большинстве случаев основной причиной, нарушающей воспроизводство стад промысловых рыб при зарегулированном стоке рек (Лапицкий, 1970; Ильина и Гордеев, 1972).
Существенным фактором, вызывающим гибель рыб, является сброс в водоемы токсических веществ, которые или непосредственно убивают икру, эмбрионов, личинок и взрослых рыб, или, разлагаясь, отнимают из воды кислород и становятся причиной заморов.
Случаи гибели рыб в послеличиночный период вследствие изменения абиотических условий редки и наблюдаются более или менее регулярно только на краю биологического ареала вида или в условиях, измененных человеком. Резкие перепады температуры могут иногда в морях вызвать массовую гибель рыб, для которых данные температуры оказываются смертельными. Естественно, что разные виды рыб по-разному реагируют на подъем или падение температуры: одни гибнут, другие выживают. При такой селективной смертности, особенно в замкнутых водоемах, может происходить коренное изменение состава ихтиофауны. Подобное явление описано, например, Бейли (Bailey, 1955) в одном озере штата Мичиган (Bass Lake), где выжили лишь немногие виды. Основной причиной гибели как в этом, так и в большинстве других случаев, однако, является не непосредственно температура, а нарушение условий дыхания. Виды, способные жить длительное время почти в анаэробных условиях, например обыкновенный карась, или дышать при помощи атмосферного воздуха, могут жить и при высоких температурах, а карась, как известно, может переносить даже длительное высыхание водоемов, зарывшись в ил (Шнитников, 1961). Вместе с тем даже незначительное повышение температуры и снижение насыщенности воды кислородом вызывают гибель ряда видов лососей. Конечно, прогнозируя возможную гибель рыб, нужно учитывать не только возможные температуры, но и состояние, в котором находится рыба.
Существенной причиной смертности рыб под воздействием абиотических условий являются заморы, возникающие в результате развития гнилостных процессов и исчезновения кислорода из воды водоемов. (Мы здесь не касаемся искусственно вызываемых заморов, возникающих в результате сброса сточных вод.) Они распространяются главным образом на лимнофильных рыб, зимующих в озерах, и иногда носят регулярный характер, особенно усиливаясь в суровые зимы. В тропиках известно много случаев, гибели рыб в результате дефицита кислорода. Так, в Индии нередко в начале периода дождей в облачную погоду резко снижается интенсивность фотосинтеза, а первые дожди смывают в сильно пересохшие водоемы много органических веществ, которые, отнимая кислород из воды, вызывают его дефицит, что приводит к массовой гибели рыб (Moitra, 1955). Летние ночные заморы, вызываемые изъятием кислорода растительностью, менее губительны, чем зимние, и отражаются главным образом на икре и молоди рыб. Однако, несомненно, что в динамике численности рыб на после личиночных этапах развития абиотические условия играют гораздо большую косвенную роль, меняя - через изменение периода нагула, величины кормового ареала, интенсивности обмена, доступности кормов - обеспеченность популяции пищей. Об этом уже говорилось выше.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'