Приспособительное значение разновозрастности полового созревания
Выше мы уже говорили, что возраст полового созревания у рыб данной популяции связан в первую очередь с достижением определенных размеров. Следовательно, чем быстрее растет рыба, тем раньше она достигнет половой зрелости. У воблы Северного Каспия Г. Н. Монастырский (1940) установил определенную линейную зависимость между средними размерами, которых достигла популяция рыб, и процентом половозрелых особей в популяции. У каспийского пузанка Caspialosa caspia (Eich.) более быстрорастущие особи становятся половозрелыми в более раннем возрасте, чем рыбы более медленно растущие (рис. 25).
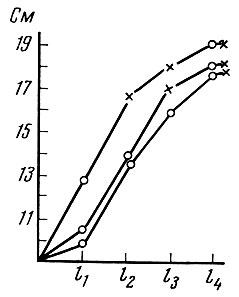
Рис. 25. Рост четырехгодовиков пузанка в зависимости от возраста первого нереста (по Замахаеву, 1949). Крестиками обозначен нерест
Аналогичная картина имеет место и у самцов Callionymus lyra (Chang Hsiao-wei, 1951) и у дальневосточных лососей (Бирман, 1953): кеты, красной и у большинства других видов рыб.
Особи разного возраста созревают примерно при одинаковой длине: пузанок при 16-18 см, Callionymus lyra 15-18 см, осенняя кета бассейна Амура при 67-70 см. Однако во всех рассмотренных случаях как у рыб с однократным нерестом (все производители кеты и самцы Callionymus lyra после первого нереста погибают), так и у рыб с повторным нерестом раньше созревающие рыбы становятся половозрелыми при несколько меньших размерах, чем более поздно созревшие (рис. 26).
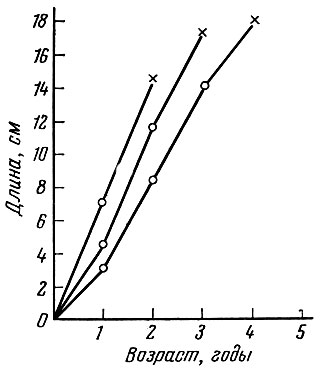
Рис. 26. Рост Callionymus lyra и время первого нереста (по Chang Hsiao-wei, 1951). Крестиками обозначен нерест
У рыб, для большинства особей которых типичен повторный нерест (например, плотва и окунь), отмечается еще одна закономерность: рыбы, растущие наиболее медленно, созревают при несколько меньших размерах, чем рыбы со средней скоростью роста (Aim, 1959). Видимо, эта особенность созревания наиболее медленно растущих особей в популяции в какой-то степени может быть связана и с механизмом образования карликовых скороспелых форм.
В известной степени схематизируя, можно, следуя Альму (1959), изобразить связь скорости роста и времени наступления половой зрелости в виде диаграммы, показанной на рис. 27.
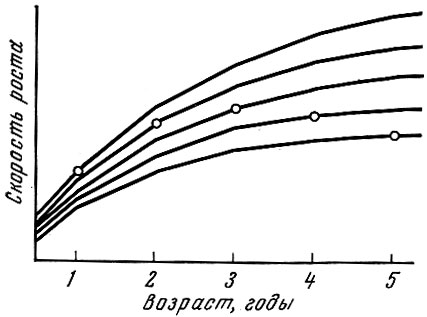
Рис. 27. Связь между скоростью роста и созреванием (по Aim, 1959). Кружками показано время наступления половой зрелости
Различный рост особей в поколении приводит к тому, что поколение созревает обычно в разном возрасте. По-видимому, лишь очень немногие виды рыб созревают все в одном возрасте. К таким рыбам могут быть отнесены некоторые виды бычков Gobiidae, некоторые живородящие карпозубые Poecilidae и, возможно, некоторые другие. Как предполагает Ю. Е. Лапин (1971), у горбуши Oncorhynchus gorbuscha (Walb.) при неблагоприятных условиях нагула часть особей созревает в возрасте 2+, причем размеры позднее созревших особей оказываются лишь немного большими, чем у рыб, созревших в возрасте 1 + . Чем сильнее у рыбы варьирует размерный ряд поколения, тем более растянутым оказывается ряд возрастного состава впервые созревающих особей. Так, например, у куринской севрюги (Державин, 1922) поколение (оба пола) созревает в возрасте от 7 до 17 лет, у амурской кеты Oncorhynchus keta (Walb.) - от 2+ до 7+ (Бирман, 1953), у атлантико-скандинавских сельдей Clupea harengus L. - от 3 до 7 (Марти, 1956), у воблы Северного Каспия - от 2 до 5 (Монастырский, 1940), у густеры Blicca bjorkna L. от 3 до 10 (Беккер, 1959). У большинства рыб самцы созревают несколько раньше самок и становятся половозрелыми обычно при несколько меньших размерах. Естественно, что возраст, в котором данное поколение входит в нерестовое стадо, зависит от ряда причин, определяющих обеспеченность пищей и рост рыб в поколении. Через изменение темпа роста меняется и возрастной состав пополнения стада. Выше мы рассмотрели, какое влияние на рост данного и смежных поколений оказывают различные поколения северокаспийского леща. У леща Северного Каспия, как показала К. А. Земская (1958), чем больше численность, тем медленнее темп роста. Эта связь численности поколения и роста отражается и на растянутости ряда впервые созревающих рыб поколений разной численности и на среднем возрасте созревания рыб разных поколений (табл. 49).
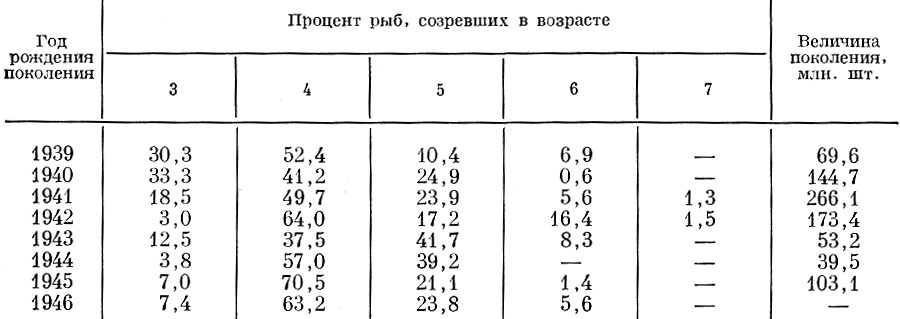
Таблица 49. Темп созревания поколений разной мощности у леща Северного Каспия (по материалам К. А. Земской)
Ряд впервые созревших особей северокаспийского леща был наиболее растянутым у более многочисленных поколений (поколения 1941 и 1942 гг.). Отчетливо выявляется отмеченное Г. Д. Поляковым (1962) взаимное влияние численности смежных поколений на ход полового созревания у рыб, их слагающих.
У кеты в связи с изменением темпа роста также меняются не только средний возраст полового созревания, но и растянутость возрастного ряда впервые нерестующих рыб. При повышении темпа роста возрастной ряд впервые нерестующих особей становится меньше, при замедлении темпа роста он растягивается, что, естественно, изменяет темп пополнения. Так, у осенней кеты Амура за 1925-1950 гг. в годы с высокой численностью время полового созревания растягивается, а в годы с низкой численностью сокращается.
Одновременно средний возраст созревания сдвигается в сторону младших возрастов (табл. 50). Это связано и с повышением плодовитости одноразмерных рыб.
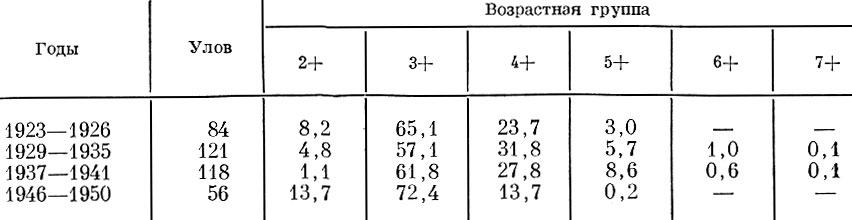
Таблица 50. Соотношение (в %) возрастных групп в нерестовом стаде осенней кеты и величина улова (в %) к многолетнему среднему (по Бирману и Леванидову, 1953)
Сходная, но несколько более сложная картина наблюдается у арктической трески; у нее в годы высокой численности время полового созревания также запаздывает и ряд впервые созревающих особей растягивается (Rollefsen, 1954).
Естественно, что рост рыб до достижения половой зрелости и возраст полового созревания определяются не только численностью поколения и величиной кормовой базы, но и продолжительностью и температурными условиями сезона нагула. Так, В. С. Кирпичников (1935) показал, что в годы потеплений корюшка Белого моря растет лучше, чем в более холодные годы.
Более южные популяции норвежской сельди растут быстрее и созревают в более раннем возрасте (Бирюков, 1956). Подобная закономерность отмечена у многих как морских, так и пресноводных рыб. Наступление половой зрелости в более раннем возрасте связано с меньшей продолжительностью жизни рыб и с упрощением возрастной структуры более южных популяций одного и того же вида.
Характер изменений режима сезона нагула на разных видах рыб отражается по-разному. Температура обычно оказывает сходное влияние на рост рыб, принадлежащих к одному и тому же или географически близким фаунистическим комплексам. Так, азовский судак и тарань лучше растут в годы с продолжительной теплой осенью. Наоборот, высокие летние температуры отражаются на росте судака и тарани отрицательно (Бойко, 1955).
Таковы основные закономерности, которым подчиняется рост рыб до достижения половой зрелости. Как видно из изложенного, основные приспособительные особенности роста рыб в этот период заключаются в томт что основные энергетические ресурсы, поступающие в организм, расходуются на увеличение размеров тела; резервных веществ в этот период в организме, как правило, накапливается очень мало (о некоторых исключениях сказано ниже).
Поэтому рыбы очень чутко реагируют на изменение обеспеченности пищей изменением темпа линейного роста и возраста полового созревания.
В этот период жизни рыбы рост является тем основным механизмом, который контролирует темп вступления пополнения в нерестовое стадо и его выживание.
Следующий короткий период в жизни рыбы, характеризующийся своей спецификой роста, - это период полового созревания. В этот период основная функция организма - обеспечение формирования половых желез, созревания половых продуктов и накопления резервных веществ в организме для поддержания обмена веществ в период нерестовой миграции и нереста, когда рыба расходует большое количество энергии, а потребление пищи сильно сокращается или прекращается полностью. В связи с этим в рассматриваемый (второй) период роста нарастание длины тела резко замедляется и начинается интенсивный процесс отложения в организме резервных веществ, в первую очередь жиров.
Правда, жирность нарастает и в первый период роста, до начала подготовки к первому нересту. Так, у разных возрастных групп леща северной части Аральского моря осенью жирность в мышцах меняется следующим образом:
| Возвраст | Количества жира в мышцах, % |
|---|---|
| 0+ | 1,4 |
| 1+ | 1,8 |
| 2+ | 2,0 |
| 3+ | 9,6 |
| 4+ | 8,3 |
| 5+ | 11,5 |
Половая зрелость наступает у аральского леща в возрасте 3+, и это связано с резким увеличением количества жира в организме. В то же время период линейного роста сокращается, и если у молоди начало быстрого роста приходится на весну, то у лещей, достигающих половой зрелости на следующий год, начало линейного роста сдвигается на середину лета и осень, а первая часть сезона нагула идет на накопление жировых резервов, необходимых для обеспечения развития половых продуктов и нереста (Константинова, 1958). У рыб, нерестующих только один раз в жизни, этот второй период роста является и последним. Этот второй период роста рыбы плохо выделяется по характеристике роста, хотя можно отметить, что для характеристики роста года созревания характерна очень большая изменчивость. Но по сравнению с предыдущим периодом роста ее изменчивость все же, как правило, снижается. Исключение представляет только те случаи, когда рост до достижения половой зрелости оказывается очень замедленным.
У рыб с повторным нерестом следующий период характеризуется снижением значения линейного роста в обменных процессах организма; основные поступающие в организм ресурсы расходуются на накопление жиров и на построение половых продуктов. Нарастание же длины происходит обычно уже после того, как в организме будет накоплен необходимый запас резервных веществ. В связи с этим у рыб старших возрастных групп закладка годового кольца и начало нового периода прироста чешуи часто приходятся на конец лета и осень, после накопления необходимого запаса резервных веществ. Это отмечено у леща Аральского моря (Константинова, 1958), судака Азовского моря (Чугунова, 1931), голубого окуня озер Северной Индианы (Ricker, 1942).
Этот период роста характеризуется у многих рыб очень значительными сезонными колебаниями массы тела и жирности, тогда как в период достижения половой зрелости сезонная динамика массы и жирности обычно незначительна. Характерной особенностью этого третьего периода роста являются очень большие различия в жирности у разных видов рыб. У рыб, у которых имеет место зимовка, связанная с длительным перерывом в питании, или у рыб, довершающих далекие миграции в теле, как известно, накапливается больше жира и сезонная динамика его более велика, чем у рыб, не совершающих далеких миграций и не зимующих. Этот период в жизни рыбы обычно характеризуется, как это отмечено у карповых и сиговых рыб В. В. Васнецовым (1947, 1953), стабилизацией линейной характеристики роста. У достигших половой зрелости рыб вырабатывается ряд регуляторных приспособлений, связанных с обеспечением нормального развития половых желез и высокого качества потомства в меняющихся условиях обеспеченности пищей.
У различных видов рыб жировые депо распределяются по-разному: у акул и тресковых основные запасы жира концентрируются в печени, у карповых и окуневых на внутренностях, у лососевых в мышцах (Шульман, 1960). Несколько могут отличаться жировые депо и у особей разного пола.
Так, у самцов трехиглой колюшки в печени содержится относительно больше жиров, чем у самок (Kilarski, 1960). Однако при концентрации основных масс жира у разных групп рыб в разных участках тела, по-видимому, у всех видов в случае голодания жир срабатывается из организма в определенном порядке и в последнюю очередь из гонад (Макарова, 1961). Иногда при голодании раньше сработки жира из гонад начинается сработка белков тела и даже некоторое сокращение длины рыб (Чугунова, Ассман, Макарова, 1961). Накопление жиров в теле достигших половой зрелости особей не только связано с обеспечением воспроизводства потомства, но имеет и некоторые другие формы приспособительного значения (см. ниже). К сожалению, приспособительное значение жирового баланса в теле рыбы как механизма, регулирующего темп воспроизводства и выживания популяции, особенно качественная сторона этого процесса, изучены недостаточно.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'