Взаимовлияние смежных поколений на рост рыб в поколении
Поколение рыб в водоеме существует не изолированно от других поколений. Чем быстрее рост рыб, чем больше разница в составе пищи у рыб разных размеров, чем больше расходятся места нагула рыб разного возраста, тем меньше одно поколение влияет на рост смежных поколений. Однако у большинства видов рыб имеет место захождение размеров рыб смежных поколений. Естественно поэтому, что особи одного поколения влияют на обеспеченность пищей особей того же размера в смежных поколениях. Быстрорастущая группа рыб данного поколения, догоняя в росте отстающую часть рыб предшествующего поколения, в случае ограниченности обеспеченности пищей оказывает на ее рост затормаживающее влияние. Естественно, что и сама эта относительно быстрорастущая часть рассматриваемого поколения начинает расти несколько хуже, вследствие чего ее догоняет в росте модальная по размерам часть поколения. Поскольку же численность данной размерной группы увеличивается за счет особей предыдущего поколения, это отразится на ее росте, что приведет к его замедлению и увеличению его изменчивости, а тем самым к усилению влияния на следующее за рассматриваемым поколение. Это взаимовлияние смежных поколений на рост особей, их слагающих, хорошо прослежено у многих рыб (Поляков, 1961, 1962; Барсуков, 1961; Le Cren, 1958; Шатуновский, 1965 и др.).
Как показал Г. Д. Поляков (1962) на материалах американских исследователей Фелина и Филлипса (Felin a. Phillips, 1948), мощное поколение калифорнийской сардины оказывает закономерное влияние на смежные поколения. У сардины, рыбы с коротким жизненным циклом, мощное поколение влияет обычно только на ближайшие предшествующее и последующее поколения, причем это влияние особенно сильно начинает сказываться с возраста 2+ (рис. 20). У леща, рыбы с более длительным жизненным циклом и более растянутым, чем у сардины, ростом и возрастным рядом впервые нерестующих особей, влияние мощных поколений сказывается не только на смежных, но и на более далеких по возрасту от мощного поколения. Так, за исключением младших возрастных групп, где, как у большинства озерных рыб, взаимовлияние смежных поколений на первом году жизни выражено слабо, намечается ясная картина влияния смежных, а иногда и не только смежных поколений леща друг на друга (рис. 21). Это хорошо прослеживается у леща Северного Каспия и Рыбинского водохранилища (рис. 22 и 23).
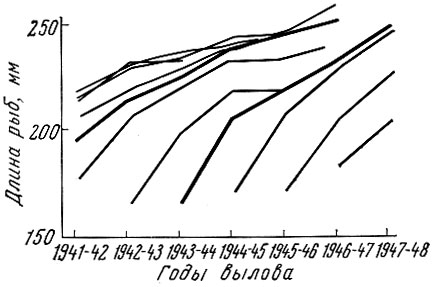
Рис. 20. Линейный рост (с годовалого возраста) поколений тихоокеанской сардины в районе Монтрей (по Полякову, 1962). Жирными линиями обозначены поколения высокой численности
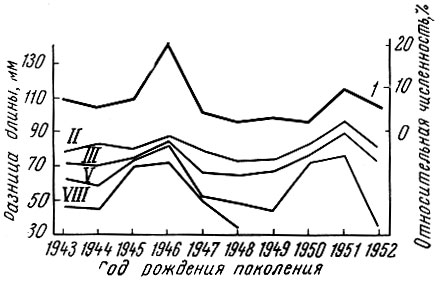
Рис. 21. Разница в длине тела рыб на год старше и на год моложе некоторых возрастных групп, представляющих разные по численности поколения рыбинского леща (по Полякову, 1962): 1 - относительная численность поколений (в % от многолетнего улова); остальные кривые - разница длины тела на год старше и на год моложе двух-, трехгодовиков и т. д. (римские цифры обозначают возрастную группу), представляющих поколения, обозначенные на горизонтальной оси графика
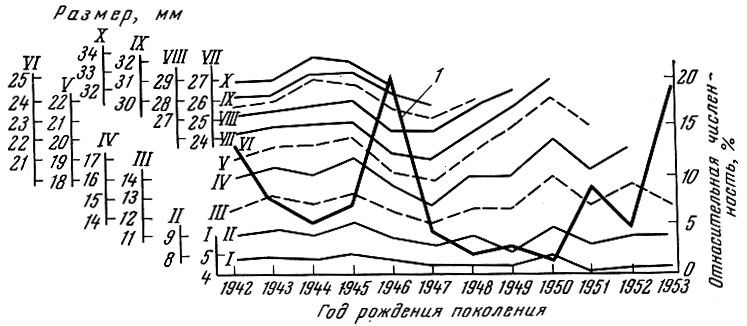
Рис. 22. Численность поколений и размер разных возрастных групп леща из Рыбинского водохранилища (по Полякову, 1962). Римскими цифрами показан возраст рыб (соответственно от I до X); 1 - относительная численность поколений (в % от многолетнего улова)
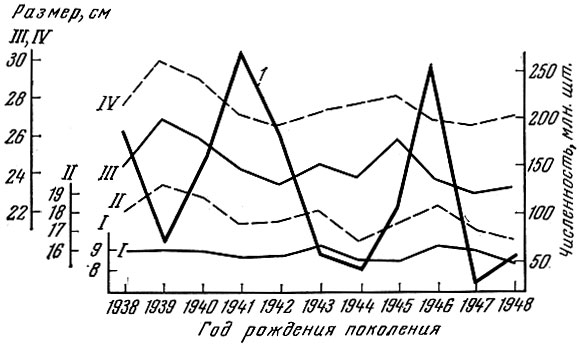
Рис. 23. Численность поколений и размеры разных возрастных групп северокаспийского леща (по данным Земской, 1959; Полякову, 1962). Условные обозначения те же, что на рис. 22
У старших возрастных групп леща (начиная с возраста 5+, 6+) также наблюдается некоторое ослабление влияния одного поколения на рост особей в смежных поколениях.
В отношении судака Рыбинского водохранилища и сига оз. Иджен (Швеция, по материалам Свердсона) В. В. Барсуков (1961) очень отчетливо показал, что высокоурожайное поколение 1946 г. (урожайное и у сига шведских озер, и у многих рыб Рыбинского водохранилища) оказало затормаживающее влияние на рост особей ряда последующих поколений и в меньшей степени сказалось на росте предшествующего ему поколения 1945 г. При этом у сига затормаживающее влияние мощного поколения на рост последующих начинает сказываться, когда рыбы достигнут 18- 20 см длины, а у судака это влияние становится заметным, когда рыбы достигнут длины около 20-25 см. У судака Рыбинского водохранилища тормозящее влияние поколения 1946 г. сказалось на росте не менее четырех поколений (1947-1950 гг.), но оно, несомненно, повлияло, хотя и в меньшей степени, на рост поколения и 1951 г. У сига оз. Иджен поколение 1946 г. оказало влияние как на последующие поколения 1947-1949 гг., так и весьма сильно повлияло на предшествующее поколение 1945 г. рождения. Такое отличие от того, что мы наблюдали у судака, объясняется более медленным ростом сига. Если у судака поколение 1945 г. благодаря быстрому росту в значительной степени успело "уйти" от тормозящего влияния поколения 1946 г., то у сига этого не получилось, и поколение 1946 г оказало сильное влияние и на темп роста поколения 1945 г.
У речной камбалы восточной части Балтийского моря взаимовлияние поколений на рост особей в смежных поколениях сказывается, начиная с сеголетков. Так, сеголетки урожайного 1957 г. оказывали заметное влияние на рост медленно росшей части годовиков 1956 г. рождения. Взаимное влияние на рост смежных поколений оказывают и рыбы старших возрастных групп.
Показатель трансгрессии кривых распределения (Т) дает формальное и заниженное представление о взаимном влиянии поколений (Шатуновский, 1965) (рис. 24).
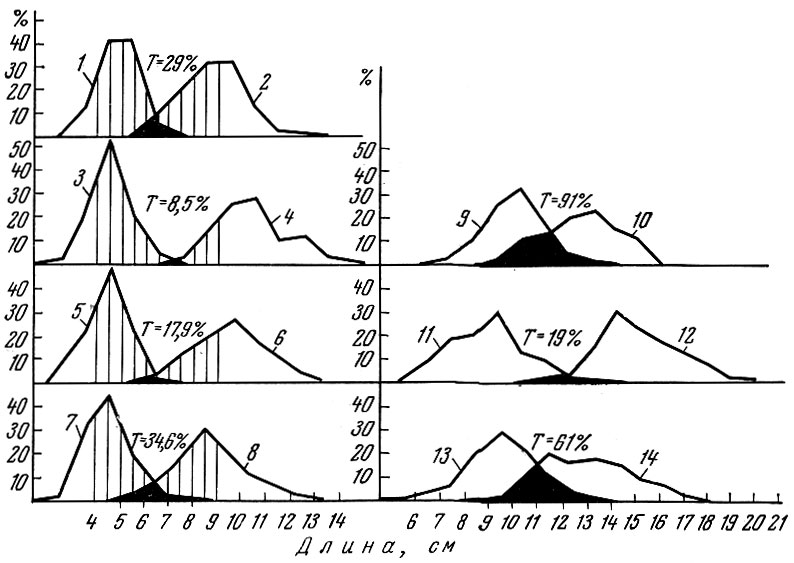
Рис. 24. Связь размерных кривых смежных поколений речной камбалы: 1 - сеголетки 1960 г. рождения; 2 - годовики 1959 г. рождения; 3 - сеголетки 1959 г. рождения; 4 - годовики 1958 г. рождения; 5 - сеголетки 1958 г. рождения; 6 - годовики 1957 г. рождения; 7 - сеголетки 1957 г. рождения; 8 - годовики 1956 г. рождения; 9 - годовики 1960 г. рождения; 10 - двухгодовики 1959 г. рождения; 11 - годовики 1959 г. рождения; 12 - двухгодовики 1958 г. рождения; 13 - годовики 1957 г. рождения; 14 - двухгодовики 1956 г. рождения
Биологический смысл такого взаимного влияния смежных поколений на рост, а тем самым и на сроки достижения половой зрелости особями этих поколений заключается в том, что мощное поколение, следующее за менее многочисленными, затормаживает темп его воспроизводства и тем самым влияет на дальнейшую обеспеченность пищей популяции. Несомненно, что это взаимовлияние поколений на рост особей, их слагающих, является одним из эффективных способов самонастройки популяции на определенный режим воспроизводства.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'