Рост и изменчивость размеров рыб
Как показал Г. Д. Поляков (1958, 1960, 1961, 1962) на примере сеголетков карпа и молоди других рыб, ухудшение обеспеченности пищей молоди тотчас сказывается не только на замедлении средней скорости роста рыб в популяции, но и на увеличении изменчивости в скорости роста. У большинства исследованных видов рыб, популяция которых находится в относительно неблагоприятных условиях, годовые приросты до достижения половой зрелости изменяются значительно сильнее, чем у рыб, достигших половой зрелости. Наоборот, в хороших условиях изменчивость величины годовых приростов у рыб до достижения половой зрелости уменьшается: все рыбы растут более или менее одинаково. У многих видов рыб плотность популяции оказывает на темп роста молоди на первом, а иногда и на втором годах жизни часто меньшее влияние, чем на последующих. Это наиболее четко отмечено у пресноводных рыб, в частности сига (Svardson, 1953), окуня Perca fluviatilis L. (Aim, 1952, 1959; Le Cren, 1958), судака (Барсуков, 1959), густеры (Беккер, 1958) и других видов. Пока молодь питается планктоном, ее рост обычно менее зависит от плотности популяции. Указанная закономерность, видимо, характерна главным образом для озерных рыб. У ряда морских рыб удается отметить связь роста с численностью и изменчивостью размеров рыб уже на первых этапах внешнего питания. Так, по Л. Н. Лисивненко (1963), коэффициент вариации линейных размеров личинок весенней салаки Рижского залива в годы с высокой обеспеченностью пищей (1955, 1957, 1959 и 1961) был много ниже (соответственно 15,3; 13,1; 11,0 и 12,6), чем в годы с низкой обеспеченностью пищей (1956, 1958 и 1960, когда он был соответственно 20,0, 26,3 и 22,7). При этом средняя длина 30-40-дневных личинок в благоприятные по обеспеченности пищей годы была 22,3 мм, а в годы неблагоприятные 19,2 мм.
У молоди трески, держащейся в открытых частях Баренцева моря (данные Т. К. Сысоевой, А. А. Дегтяревой и И. Я. Пономаренко), также установлена связь обеспеченности пищей с ростом, жирностью и изменчивостью линейных размеров. У проходных и полупроходных рыб изменчивость роста молоди в реке и в полойный период значительно выше, чем у рыб, скатившихся в море. Так, М. Н. Лишев и Е. Я. Римш (1961) показали, что изменчивость линейного и весового роста балтийского лосося наиболее велика в первый год речной жизни и первый год выхода в море (рис. 19). В эти же периоды наименьшей оказывается и относительная скорость роста.
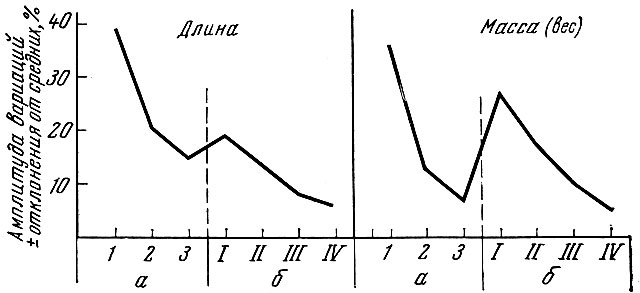
Рис. 19. Амплитуда вариаций длины и массы лосося р. Даугавы по возрастам (величина отклонений, % от соответствующих средних) (по Лишевуи Римшу, 1961): а - пестрятки и смолты; б - жизнь в море до нереста; 1, 2, 3 - годы жизни в реке; I, II, III, IV - годы жизни в море
Сходная картина наблюдается при сравнении величины изменчивости размеров молоди воблы и леща в полоях с размерами молоди воблы и леща в морской период жизни. Изменчивость молоди воблы и леща в большинстве полоев много выше, чем на местах нагула в море перед устьями Волги. В рыбхозах дельты Волги у молоди воблы абсолютный и относительный прирост находятся в обратной связи с изменчивостью роста и асимметрией кривой размерного состава (Поляков, 1960).
Характер изменчивости молоди нерки в период жизни в озерах отражается на структуре ее косяков в море (Крогиус, 1960). Сказанное подтверждает изложенные выше данные о разнице в обеспеченности пищей разных возрастных категорий у видов рыб, принадлежащих к различным экологическим группам. Расхождение в размерах рыб при ухудшении обеспеченности пищей есть не только приспособление к расширению спектра питания, а тем самым и кормовой базы популяции, но это расхождение приводит к созреванию поколения в разном возрасте; при ухудшении обеспеченности пищей это является благоприятным, так как создает большую равномерность пополнения.
Расхождение в скорости роста особей в поколении часто связано и с усилением разницы в росте самцов и самок. Так, в наиболее плохих по кормности условиях у обыкновенного карася Carassius carassius (L.) наблюдается наибольшее расхождение в росте самцов и самок. Самцы гораздо сильнее отстают в росте от самок, чем в обычных средних условиях питания (Беккер, 1959). Наоборот, при улучшении обеспеченности пищей разница в росте самцов и самок сглаживается. Более сильное в плохих условиях питания затормаживание роста самцов, чем самок, позволяет популяции и в ухудшившихся условиях обеспеченности пищей в меньшей степени снизить интенсивность воспроизводства, чем если бы рост обоих полов затормаживался в одинаковой степени.
Каков механизм, приводящий к дивергенции в росте рыб, перешедших к активному питанию, пока точно не известно. У молоди окуня расхождение в росте у особей в помете одной самки начинается уже с момента выхода из икры. Дальше дифференциация в размерах связана с особенностями питания. Более крупные особи - 10,4-12,7 мм длины, находящиеся на VI-VII этапах развития, переходят на питание наиболее мелкими особями 6,8-10,3 мм длины, находящимися на IV-V этапах развития, которые питаются планктоном. В дальнейшем разница в размерах особей, питающихся разным кормом, усиливается. Разные по характеру питания группы молоди окуня отличаются и своим поведением (Ильина, 1970). При разной плотности популяции и разной обеспеченности пищей меняется поведение рыб в группировке. При недостатке пищи усиливается разница в поведении разных особей, при повышении обеспеченности пищей эта разница в поведении сглаживается (Kawanabe, 1958 и др.).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'