Соотношение весового и линейного роста
Если темп линейного роста до достижения половой зрелости обеспечивает рыбе быстрый выход из-под воздействия хищников при сравнительно малом расходовании кормов, то после достижения половой зрелости он снижается, но резко возрастает темп весового прироста массы. Так, у леща Волго-Каспийского района линейная характеристика роста после достижения половой зрелости снижается с 6,4 до 3,9, а характеристика весового роста возрастает с 12 до 16. У сазана Арала линейная характеристика роста по достижении половой зрелости снижается с 6,6 до 5,2, а весовая возрастает с 27 до 30. У сазана Арала хорошо выделяется и третий период - период старения, когда характеристика весового роста падает с 43,1 до 25,0. У беломорской корюшки линейная характеристика роста снижается по достижении половой зрелости с 4,7 до 3,4, а весовая возрастает с 15,3 до 25,7. У сельди залива Анива наблюдается сходная картина: темп линейного роста снижается по достижении половой зрелости, а весовой рост возрастает. Высокий весовой рост сохраняется у сельди до возраста 7+-8+, когда, видимо, у этой сельди начинается период старения.
У многих рыб линейный рост более интенсивен до достижения половой зрелости, а максимальные абсолютные приросты массы тела наблюдаются у половозрелых рыб. Старение у многих рыб с повторным нерестом, как это отмечено по изменению характеристики роста у сазана, выражается в переломе кривой нарастания массы в сторону снижения. Этот перелом у разных популяций одного вида и в пределах одной популяции в разные по обеспеченности пищей годы может приходиться на разные возрасты и связан с возрастом достижения половой зрелости. Л. С. Бердичевский (1964), пользуясь формулой
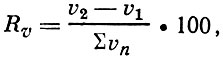 (6)
(6)где:
Rυ - рост массы;
υ2 и υ1 - годовые приросты массы;
Συn - сумма годовых приростов,
показал, что максимальное нарастание массы тела приходится далеко не всегда на первые половозрелые группы.
Л. С. Бердичевский правильно указывает, что расчисление массы в процентах к массе за предыдущий год не дает действительного представления об абсолютной величине нарастания массы тела. Но я не могу согласиться с Л. С. Бердичевским, что для анализа роста массы рыбы нельзя пользоваться характеристикой роста, введенной В. В. Васнецовым (1934). Я считаю, что расчисление темпа роста по показателям прошлого года может быть использовано для решения ряда как биологических вопросов, так и практических задач. Приведенные выше примеры анализа весового роста говорят о применимости характеристики роста для выявления качественно специфичных периодов индивидуального роста рыбы как весового, так и линейного. Л. С. Бердичевский (1964) упрощает явление, говоря, что между линейным и весовым ростом рыбы существует обратная связь. На самом деле в одни периоды онтогенеза эта связь прямая, в другие - обратная.
Быстрое наращивание массы тела в период половой зрелости есть важное приспособление, обеспечивающее значительное увеличение плодовитости стада, ибо, как известно, плодовитость у рыб коррелирует в большей степени с массой тела рыбы, чем с длиной или возрастом (см., например, Спановская, Григораш и Лягина, 1963).
Линейный рост рыбы, как правило, необратимый процесс, лишь при метаморфозе у личинок миноги - пескороек при превращении во взрослую миногу несколько уменьшаются линейные размеры. Небольшое уменьшение длины отмечено в условиях сильного голодания у сазана (Чугунова, Ассман и Макарова, 1961). Весовой же рост подвержен очень большим сезонным колебаниям (см. ниже). У большинства бореальных видов рыб, особенно тех, у которых имеется состояние зимовки, максимальная биомасса как отдельных особей, так и популяций в целом, приходится на осень, а минимальная - на посленерестовое время. У арктических и тропических рыб разница в биомассе в разные сезоны года выражена обычно значительно слабее.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'