Различия в плодовитости разных популяций одного и того же вида
Популяции одного вида, живущие в разных условиях, естественно различаются по плодовитости. Разница между плодовитостью популяций одного вида, а также близких видов, обладающих сходной морфо-экологи-ческой защищенностью от врагов, отражает, в каких условиях обеспеченности пищей и при каком прессе хищников популяция живет.
Чем сильнее различаются условия, в которых живут разные популяции вида, тем больше различается их плодовитость. Так, у популяций сельди Clupea harengus L., населяющих воды северной части Тихого океана и Северной Атлантики, наблюдаются значительные различия в плодовитости (рис. 15).
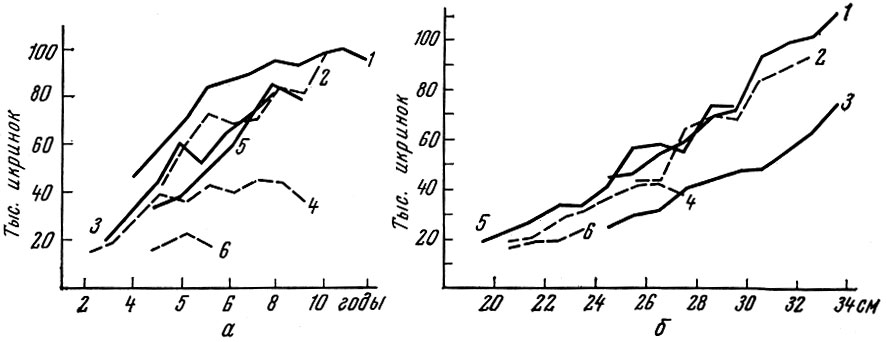
Рис. 15. Абсолютная плодовитость разных стад сельди Clupea harengus L. по возрастным (а) и размерным (б) группам (по материалам ТИНРО, Ловёстофтской станции, данным Наумова, 1956 и собственным): 1 - сахалинская; 2 - атлантическо-скандинавская; 3 - Бристоль (Тихий океан); 4 - Северное море; 5 - залив Уала (Тихий океан); 6 - мурманская
Особенно отчетливо в Тихом океане и менее четко в Северной Атлантике обнаруживается увеличение плодовитости одноразмерных и одновозрастных сельдей в направлении с севера на юг. Северные популяции, как правило, менее плодовиты, чем южные. При этом у южных популяций более быстрый рост и более раннее половое созревание не всегда связаны с сокращением продолжительности жизни. Особенно отчетливо это заметно на примере сельди Южного Приморья (Амброз, 1931).
Отсутствие сокращения продолжительности жизни в связи с более ранней половой зрелостью приводит к очень значительному возрастанию популяционной плодовитости. Плодовитость сельдей восточной части Тихого океана (Бристольский залив, наши данные) лишь немного выше плодовитости соответствующих размерных групп сельди Южного Сахалина.
Естественно, что плодовитость каждой популяции не остается постоянной из года в год, как это показано нами на примере сельди залива Уала и широкопадинской сельди Южного Сахалина.
Представляет интерес плодовитость сельди оз. Тоннай на восточном побережье Сахалина (Пробатов, Фролов, 1951): по сравнению с другими сельдями Тихого океана это - скороспелая мелкая сельдь с малой плодовитостью.
Значительно более сложная картина наблюдается у популяций сельдей Северной Атлантики. Очень значительные различия в плодовитости наблюдаются у популяций, различающихся по времени нереста - весенне-, летне- и осенненерестующих, хотя, например, у балтийской салаки эти стада и не являются полностью обособленными (Sjöblom, 1960; Анохина, 1969; Чепурнов, 1967). Весенненерестующие сельди обычно менее плодовиты, чем осенненерестующие. Осенняя салака Рижского залива, обычно, значительно более плодовита, чем весенняя. Более плодовитыми оказываются как рыбы одинаковой длины, так и возраста (табл. 38).
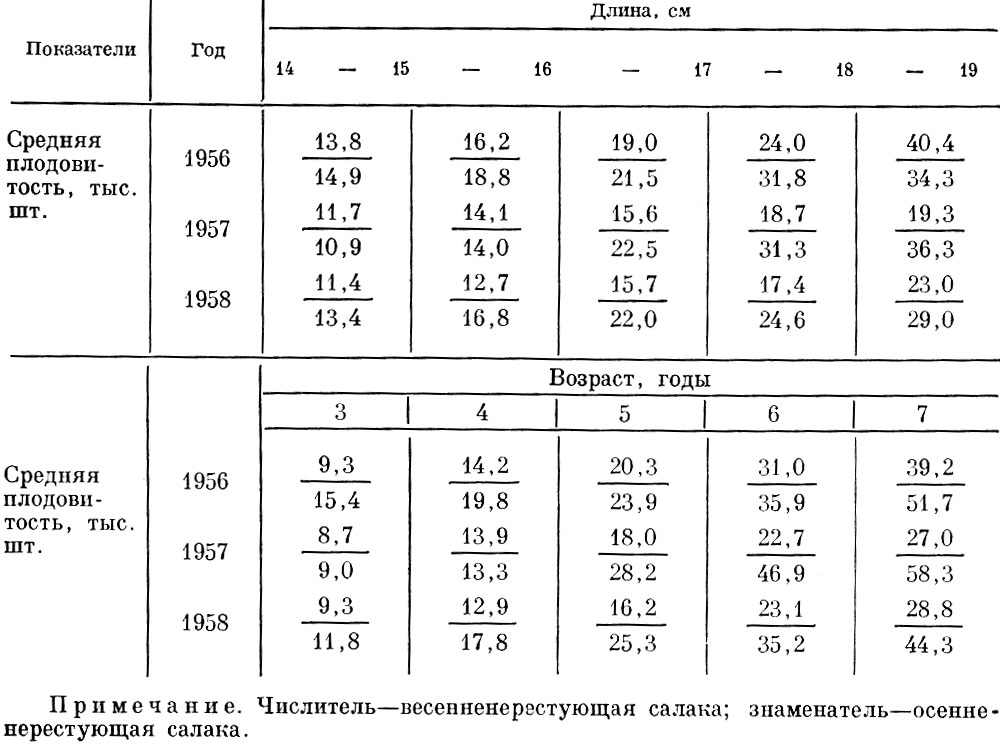
Таблица 38. Средняя индивидуальная плодоритость весенне- и осенненерестующей салаки рижского залива (по нохиной, 1969)
Отдельные отклонения наблюдаются главным образом у младших возрастных групп. Они все приходятся на 1957 г., что, несомненно, связано с низкой обеспеченностью пищей осенней салаки, нерестовавшей в 1957 г. У сельдей Белого моря гораздо плодовитее оказываются летненерестующие сельди. Как показал С. Г. Соин (1963, 1964), эти сельди размножаются в условиях значительно большей активности хищников (главным образом трехиглой колюшки) и гибель икры у них гораздо больше. Плодовитость летненерестующих сельдей Белого моря повышается при уменьшении количества желтка в икринке. Но так как летненерестующая сельдь развивается при большем развитии кормовой базы, то значительного снижения обеспеченности пищей молоди это сокращение запаса желтка, видимо, не вызывает. Сельди северной части Тихого океана плодовитее сельдей Северной Атлантики. Эта общая закономерность отмечается у популяций как одного вида, так и близких видов. Как в морских, так и в пресных водах Дальнего Востока рыбы обычно плодовитее рыб Северной Атлантики (табл. 39), а также, как будет показано ниже, у них сильнее развиты и другие приспособления против интенсивного воздействия хищников.

Таблица 39. Абсолютная плодовитость подвидов одного и того же вида рыб в бассейне тихого и атлантического океанов (в тыс. шт. икринок)
Способы достижения большей плодовитости у рыб различны; это - или более быстрый рост и достижение половой зрелости рыбами больших размеров при одном и том же возрасте (многие рыбы Тихого океана растут быстрее, чем атлантические, например мойва), или большая плодовитость достигается благодаря уменьшению размеров икринок (амурская минога, калуга). В некоторых случаях это достигается вследствие большего объема, который занимает зрелый яичник в полости тела рыбы, как это наблюдается у жилой красной по сравнению с проходной (Смирнов, 1959). Может также иметь место и большая плотность желтка икринки при несколько меньших ее размерах. Таким образом достигается повышенная концентрация кормов для эмбриона в период питания желтком.
Весьма существенны различия плодовитости и у разных популяций лососевых рыб. Так, например, по данным К. А. Савваитовой (1962), плодовитость проходной формы Salvelinus alpinus (L.) Камчатки несколько выше, чем у жилых форм. Менее плодовитой оказывается озерно-реч-ная и наименее плодовита, по-видимому, озерная форма. У различных популяций озерных гольцов плодовитость различна, причем плодовитость так называемой лудожной палии обычно выше, чем ямной (Смирнов, 1933).
Большая плодовитость проходных и полупроходных форм отмечена и у многих других рыб. Так, плодовитость полупроходных форм плотвы Rutilus rutilus (L.) обычно намного выше, чем у жилых. У некоторых тугорослых форм плотвы, живущих в условиях низкой обеспеченности пищей и при высоком прессе хищников, например у плотвы из озер по Северному Узбою, наблюдается половое созревание у более мелких рыб и повышение плодовитости. (Подробный обзор динамики плодовитости у разных форм плотвы дан Спановской, Григораш и Лягиной, 1963.)
У леща Abramis brama L. характер динамики плодовитости у отдельных популяций во многом близок к наблюдаемому у плотвы, а именно: полупроходные формы оказываются плодовитее жилых, в ряде мест образовались карликовые популяции с более ранним созреванием и высокой относительной плодовитостью. Южные формы обычно плодовитее северных. Однако у леща механизм регуляции плодовитости у северных и южных популяций несколько различен. Если у плотвы повышение плодовитости более южных популяций достигается путем ускорения роста особи и иногда увеличения относительной плодовитости, то у леща, кроме того, в южных районах, как отмечено выше, часто появляется порционность нереста (Мейен, 1940).
Значительная разница в плодовитости наблюдается у разных популяций щуки и обыкновенного окуня (Попова, 1971). Это связано в первую очередь с различной интенсивностью воздействия хищников, к которому популяция приспособлена. Так, в Аральском море на нерестилищах в зарослях прибрежной зоны пресс хищников, видимо, меньше, чем в Каспии, а кормовые условия на Арале для молоди хуже, чем на Каспии. В итоге мы наблюдаем значительную разницу в плодовитости одних и тех же видов в Арале и Каспии (табл. 40).
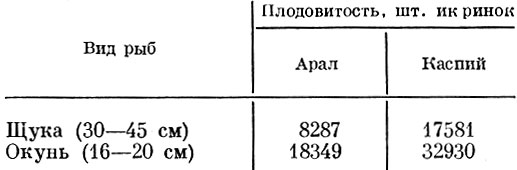
Таблица 40. Плодовитость щуки и окуня в арале и каспии (в приустьевых районах) (по Летичевскому, 1946)
Темп роста и время наступления половой зрелости у щуки и окуня в Арале и Каспии более или менее сходны.
Разница в плодовитости разных популяций одного вида наблюдается часто и в пределах одного водоема. Так, у окуня, населяющего отдельные соры Байкала, плодовитость сильно различается. Даже в пределах одного сора, например Посольского, плодовитость окуней двух рас - крупной и мелкой - очень различна (Евтюхова-Рекстин, 1962).
У щуки также удается подметить существенную разницу в плодовитости на разных участках одной и той же реки. Так, по данным А. В. Лукина и А. Н. Штейнфельд (1949), щука Средней Волги значительно менее плодовита, чем Нижней и Северного Каспия.
Морским рыбам других групп присущи те же закономерности, которые отмечены выше для сельди, а именно: почти у всех проанализированных видов морских рыб южные популяции оказываются плодовитее северных. Это отмечается как у арктических по происхождению рыб, так и у более южных. Так, из популяций наваги Eleginus navaga Pall, наиболее плодовита навага южной части Белого моря. Мезенская навага и навага Карской губы менее плодовиты (Анухина, 1962). Сходная картина наблюдается и у многих тихоокеанских Cottidae: более южные популяции одних и тех же видов Triglops, Myoxocephalus и других плодовитее северных, но запас желтка в икринке у них много меньше (Паракецов, 1962). У морской камбалы более южные и соответственно более быстрорастущие популяции тоже обладают большей плодовитостью (табл. 41).

Таблица 41. Плодовитость морской камбалы (по Bagenal, 1963)
Несколько более сложная картина наблюдается у речной камбалы. Средняя плодовитость Pleuronectes flesus L. длиной 28-30 см приведена ниже (Шатуновский, 1964).
Плодовитость одноразмерных морских камбал южных популяций более чем в 2 раза выше, чем северных.
| Район | Абсолютная плодовитость, тыс. шт. икринок |
|---|---|
| Белое море | 1250 |
| Восточная Балтика | 680 |
| Гданьский залив | 460 |
| Аркона-Борнхольм | 600 |
| Кильская бухта | 780 |
| Юго-восточная часто Северного моря | 540 |
Самой плодовитой из исследованных оказалась беломорская популяция речной камбалы, что связано со специфическими неблагоприятными условиями этого водоема. Камбалы, населяющие Балтийское море и юго-восточную часть Северного моря, по плодовитости различаются довольно слабо.
К сожалению, у нас нет данных о плодовитости самых южных популяций северных видов; имеющиеся косвенные данные показывают, что на южной границе ареалов у холодноводных форм часто образуются карликовые скороспелые формы с мелкой икрой и высокой относительной плодовитостью.
У антарктических Notothenia larseni Lönnb. и N. nudifrons Lönnb. в северном более теплом районе - у о-ва Южная Георгия - плодовитость много выше, чем в более холодном южном районе у Оркнейских и Шетландских островов. Так, у N. larseni Lonnb. длиной 17-18см в северном районе средняя плодовитость 7360 икринок, а в южном - 3094. У N. nudifrons Lönnb. длиной 14-15 см в северном районе средняя плодовитость - 4087 икринок, а в южном 1665 (Пермитин и Сильянова, 1971).
И у морских рыб субтропических по своему происхождению более южные популяции в северном полушарии также оказываются обычно плодовитее. Так, популяция малого желтого окуня Pseudosciaena polyactis Bleek., распространенная в северной части Желтого моря, несколько менее плодовита, чем распространенная в южной части Желтого моря (табл. 42).
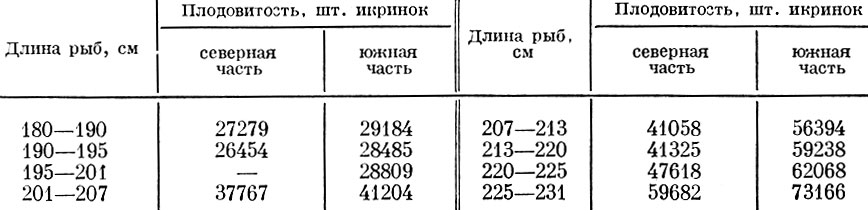
Таблица 42. Плодовитость малого желтого окуня в северной и южной частях Желтого моря (по Лю Сяо-шунь, 1962)
К сожалению, закономерности динамики популяционной плодовитости рыб экваториальной и тропической зон изучены еще очень плохо. Несомненно, что у рыб в тропической зоне механизмы, регулирующие плодовитость популяции, разнообразнее. В дополнение к механизмам, регулирующим плодовитость рыб в умеренных и высоких широтах, включаются еще механизмы, связанные с порционностью.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'