Изменение плодовитости в пределах одной популяции
Как было уже видно из анализа возрастных изменений плодовитости, она не остается постоянной у одной и той же популяции, а приспособительно меняется в связи с изменениями обеспеченности популяции пищей в разные годы. Естественно, эти изменения не бесконечны и не выходят за пределы видовой специфики, ограничиваясь определенными пределами. Увеличение или уменьшение обеспеченности пищей вызывает изменение плодовитости лишь до определенной величины, и дальнейшее изменение условий уже не влияет на изменение плодовитости. В связи с изменением условий жизни меняется как плодовитость одноразмерных особей, так и воспроизводительная способность популяции в целом, меняется соответственно и качество половых продуктов, в первую очередь размеры икринок и запас желтка в икре. Амплитуда изменений плодовитости у особей одного и того же размера или возраста может быть весьма велика, изменяясь иногда у одноразмерных рыб почти в 2 раза.
У рыб одного и того же размера или одного и того же возраста в разные годы плодовитость может быть весьма различной. Так, например, у сельди Clupea harengus (L.) юго-западной части Северного моря, по данным Hickling (1940), неопубликованным данным Cushing и данным Baxter (1958), у рыб одного итого же размера в 1956 и 1957 гг. плодовитость была много выше, чем у рыб того же размера в 1933 г. (табл. 21).
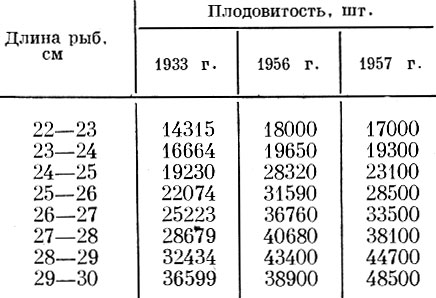
Таблица 21. Плодовитость сельди юго-западной части северного моря (по Hickling, 1940; материалам Cushing; Baxter, 1958)
По-видимому, у сельди южной части Северного моря, судя по данным голландских исследователей, реакция регуляторных механизмов с 1955 г. достигла предела и дальнейшее увеличение плодовитости в связи с улучшением обеспеченности популяции пищей в результате интенсивного промысла оказывается уже невозможным (Никольский и Белянина, 1959).
Изменение плодовитости одноразмерных особей широкопадинской сельди по годам отмечено также И. А. Пискуновым (1952) (табл. 22).
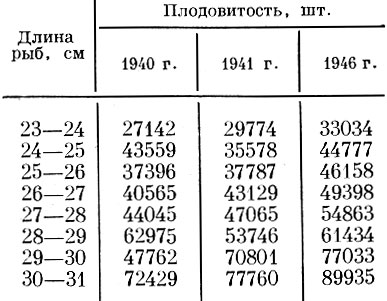
Таблица 22. Плодовитость широкопадинской сельди сахалина (по Пискунову, 1952)
Как у сельди южной части Северного моря, так и у широкопадинской сельди увеличение плодовитости одноразмерных особей связано с уменьшением численности, а тем самым с улучшением обеспеченности пищей.
Значительно изменяется плодовитость по годам и у сельдей залива Уала (Камчатка) (табл. 23).
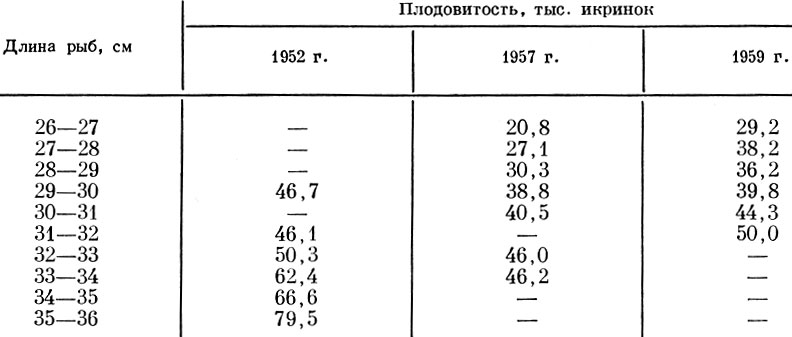
Таблица 23. Изменение плодовитости сельди залива Уала
У сельди сахалино-хоккайдского стада А. Н. Пробатов и И. Г. Фридлянд (1957) также отмечают значительные изменения плодовитости за разные годы у рыб, принадлежащих к одним и тем же возрастным группам (табл. 24).
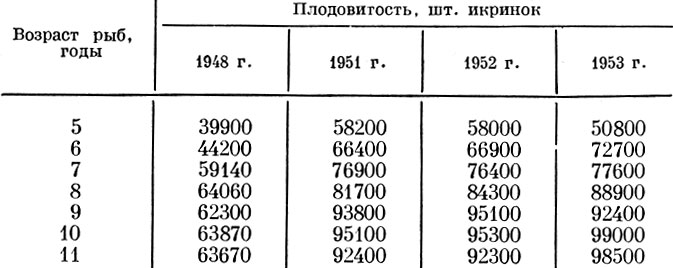
Таблица 24. Плодовитость сельдей сахалино-хоккайдского стада
Стадо сахалино-хоккайдской сельди в результате резкого снижения численности становится все меньше и меньше. Разреживание стада приводит к улучшению обеспеченности пищей и увеличению плодовитости одновозрастных рыб. Интересно, что появление несколько более многочисленного поколения 1948 г. рождения (пятилетие рыбы в 1953 г.) тотчас сказалось на снижении плодовитости этого поколения.
При сопоставлении плодовитости сахалино-хоккайдской сельди одного и того же возраста, но разного размера, как правило, плодовитее оказывались более крупные рыбы. Значительно менее четкая картина получается при сравнении плодовитости одноразмерных рыб, но разного возраста (табл. 25). Хотя и в этом случае плодовитость рыб старшего возраста увеличивается, но эта закономерность прослеживается далеко не всегда. Видимо, нарушения этой общей закономерности связаны с разной численностью поколений разных годов рождения.
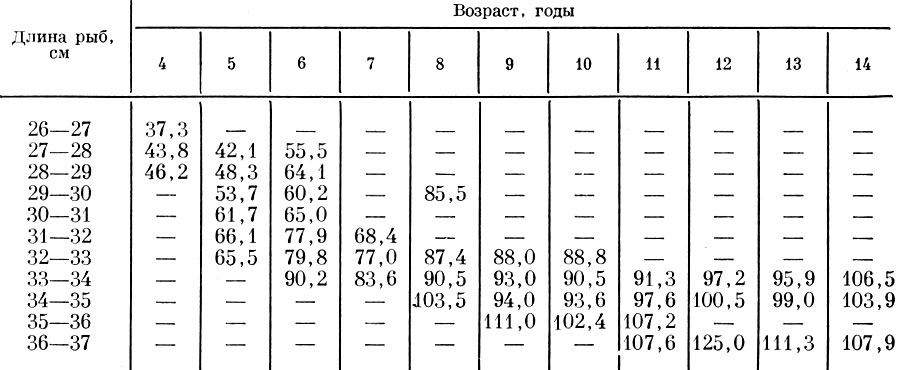
Таблица 25. Изменения плодовитости сахалино-хоккайдской сельди у рыб разного размера и возраста по сборам 1952 и 1953 гг. (в тыс. шт. икринок) (по Пробатову и Фридлянд, 1957)
У салаки залива Пярну, по данным Л. А. Раннак (1958а), плодовитость рыб в одни годы (например, 1952 и 1955) оказывается выше, в другие (1951 и 1953 ) ниже (табл. 26). Сильнее всего плодовитость колеблется у рыб младших возрастов. У салаки также выражена прямая связь плодовитости с величиной линейного прироста и прироста массы (рис. 12).
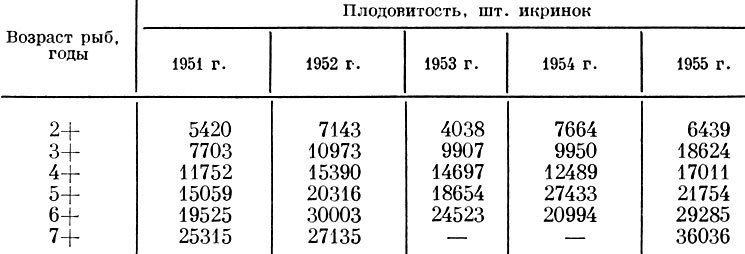
Таблица 26. Средняя плодовитость весенненерестующей салаки разных возрастных групп из залива пярну по годам (по Раннак, 1958)
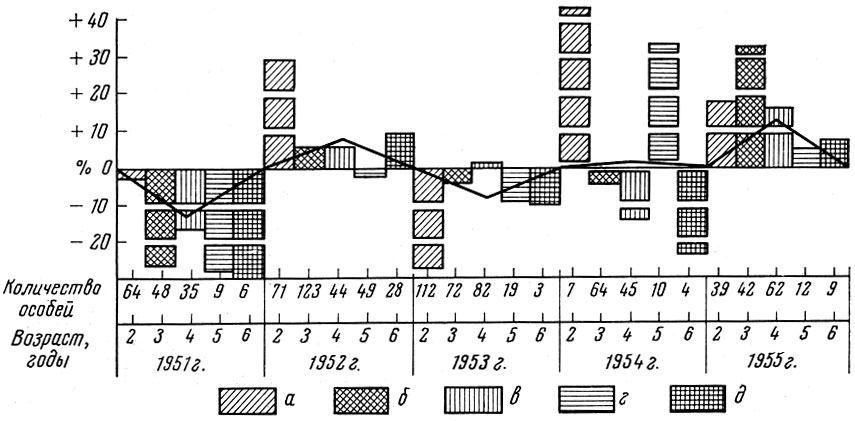
Рис.12. Отклонение плодовитости салаки (в %) от средней и прирост ее массы и длины по годам (по Раннак, 1958а). Плодовитость рыб в возрасте: а - двух лет; б - трех лет; в - четырех лет; г - пяти лет; д - шести лет
Существенные различия в пределах одного поколения наблюдаются в плодовитости одноразмерных и одновозрастных сельдей разной жирности, т. е. нагуливавшихся в разных условиях. Как показала Л. Е. Анохина (1959), более жирные самки балтийской салаки как весенне-, так и осенненерестующие, обладают большей как индивидуальной (абсолютной), так и относительной плодовитостью (табл. 27). Так как у сельди рыбы старших возрастов обычно жирнее рыб младших возрастных групп, то это делает понятным отмеченную А. Н. Пробатовым и И. Г. Фридлянд (1957) закономерность - большую плодовитость одноразмерных рыб старшего возраста. Кроме того, есть определенная связь между величиной плодовитости рыбы и тем, впервые или повторно она нерестует. Как показано Л. Е. Анохиной (1962) на примере мелкой сельди Онежского залива Белого моря, у рыб младших и старших возрастных групп плодовитее оказываются впервые нерестующие особи. У рыб средних возрастных групп эта закономерность выражена менее четко.
Как видно из приведенных цифр, более жирные рыбы, как правило, оказываются и более плодовитыми; отдельные исключения не меняют общей закономерности. Однако, как показали М. Н. Кривобок и О. И. Тарковская (1962), у салаки связь между плодовитостью и жирностью носит не прямой, а опосредованный характер.
Существенно изменилась плодовитость салаки, пересаженной в Аральское море. В связи с резким увеличением обеспеченности пищей ее плодовитость очень сильно возросла по сравнению с плодовитостью салаки Балтики. Так, у особей в возрасте 4 лет средняя плодовитость у аральской салаки составляла 60959, у весенней салаки Рижского залива - 14642, из Вислинского залива - 28000 икринок (Быков, 1962). Однако при сравнении одноразмерных рыб из Рижского залива и Арала разница оказывается не столь уж большой. Так, плодовитость весенней салаки Рижского залива длиной 18 см колеблется от 12 до 18, у осенней - от 30,3 до 47,1, а у салаки из Арала - от 8,5 до 32,5 тыс. икринок. Таким образом, увеличение плодовитости у салаки из Арала достигается не увеличением числа икринок у одноразмерных особей, а благодаря более быстрому росту и большим размерам одновозрастных рыб.
В отношении черноморской хамсы P.M. Павловская (1963) показала, что чем больше прирост массы у годовика, тем выше плодовитость особи (рис. 13).
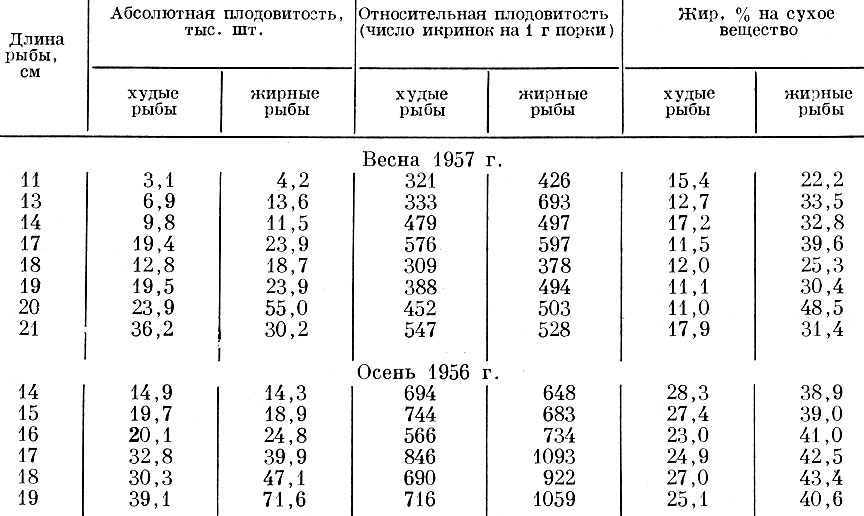
Таблица 27. Абсолютная и относительная плодовитость и жирность весенне- и осенненерестующей салаки рижского залива у рыб разного размера (по Анохиной, 1959)
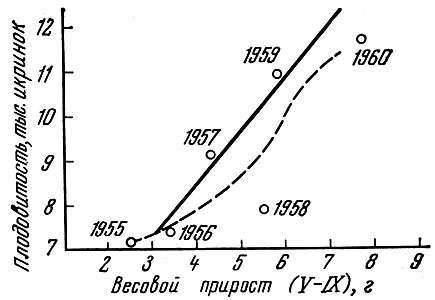
Рис. 13. Прирост массы и плодовитость годовиков черноморской хамсы (по Павловской, 1963); пунктирная линия проведена нами
Популяционная плодовитость у сельдей, а возможно и у ряда других рыб в разные годы может изменяться также вследствие пропуска частью особей нерестового сезона. Как установлено Ю. И. Чепраковой (1966) в отношении весенней Кандалакшской сельди, в неблагоприятные для нагула годы часть особей в популяции, а именно более худые рыбы не созревают к следующей весне и пропускают один нерестовый сезон, тем самым снижая плодовитость популяции. У сигов озер Лапландии и гольцов Камчатки пропуск нерестового сезона - закономерное явление (Решетников 1966; Савваитова, 1963). То же наблюдается у леща озер Карелии, т. е. на северной границе области его распространения. В то же время у леща средней полосы России нерест, как правило, каждый год, но единовременный, а в южной части его ареала у многих популяций наблюдается порционное икрометание (Кошелев, 1966).
У лососевых рыб плодовитость также сильно меняется у рыб одного размера и одного возраста в разные годы. Так, у горбуши Oncorhynchus gorbuscha (Walb.) бассейна Амура в четные годы, когда нерестовое стадо имеет более высокую численность, плодовитость рыб одного и того же размера ниже, чем в нечетные годы (Никольский, 1953).
У камчатской горбуши также наблюдается большая разница в плодовитости одноразмерных особей "урожайных" и "неурожайных" лет (Грачев, 1971).
Различаются по плодовитости одноразмерные особи разных годов рождения и у кеты (Белянина, 1963). Различия в плодовитости одноразмерных рыб наблюдаются и у семги - Salmo salar L. Так, у семги в р. Печоре плодовитость одноразмерных рыб в 1953 г. была выше, чем в 1957 г.; возможно это связано с разной численностью нерестовых стад в эти годы (Владимирская, 1960).
У семги повторно нерестующие особи, видимо, несколько плодовитее впервые нерестующих. У севанской форели-боджака отмечена прямая связь относительной плодовитости и величины коэффициента упитанности (Смолей, 1966).
Очень сильно меняется плодовитость в разные годы и у сигов. Так, по данным Б. Г. Иоганзена и А. Н. Петкевича (1958), плодовитость обского сырка Coregonus peled (Gmel.) в возрасте 4+ и 5+ за 1947-1953 гг. менялась следующим образом (табл. 28).

Таблица 28. Средняя плодовитость обского сырка Coregonus peled (Gmel.)
Таким образом, плодовитость одновозрастных особей сырка может различаться в разные годы более чем в 2 раза.
Менее значительны колебания плодовитости по годам у муксуна Coregonus muksun (Pall.), но они также достаточно четко выражены. По данным Б. Г. Иоганзена и А. Н. Петкевича (1958), у рыб в возрасте 10+ и 11+ плодовитость менялась по годам следующим образом (табл. 29).

Таблица 29. Изменение плодовитости муксуна в р. Оби (по Иоганзену и Петкевичу, 1958)
Плодовитость карповых и вьюновых рыб в пределах одной популяции также значительно колеблется в разные годы у одноразмерных и одновозрастных рыб. Так, у северокаспийской воблы, как показала Н. И. Чугунова (1951), в результате изменения условий питания плодовитость резко меняется по годам; 1946 г. по кормовым условиям для воблы был очень благоприятным и это сказалось на повышении плодовитости одноразмерных рыб.
Ниже приведена динамика плодовитости северокаспийской воблы длиной 17,5 - 18,5 см.
| Год 1911, 1912 | Плодовитость, шт |
| 1931 | 25400 |
| 1946 | 38500 |
| 1947 | 22800 |
| 1948 | 27700 |
Существенно изменяется плодовитость и речных форм плотвы в условиях образования водохранилищ. Так, сравнение плодовитости плотвы в первый (1960) и второй (1961) годы существования Можайского водохранилища показывает, что резкое увеличение обеспеченности пищей летом 1960 г. в связи с заполнением водохранилища привело к достижению плотвой половой зрелости в более раннем возрасте, повышению абсолютной и относительной плодовитости одновозрастных рыб (табл. 30), но вместе с тем к сокращению числа нерестов в течение жизни и в итоге к уменьшению общего количества икринок, выметываемых особью (табл. 31).
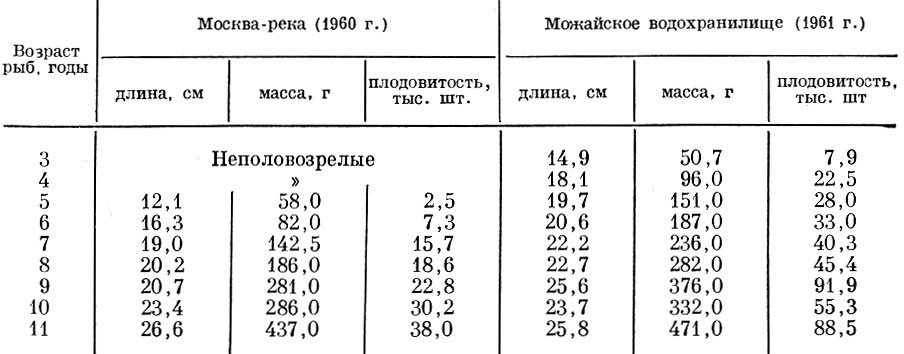
Таблица 30. Изменение плодовитости плотвы в можайском водохранилище по сравнению с ее плодовитостью в москве-реке (по данным Спановской, Григораш и Лягиной, 1965)
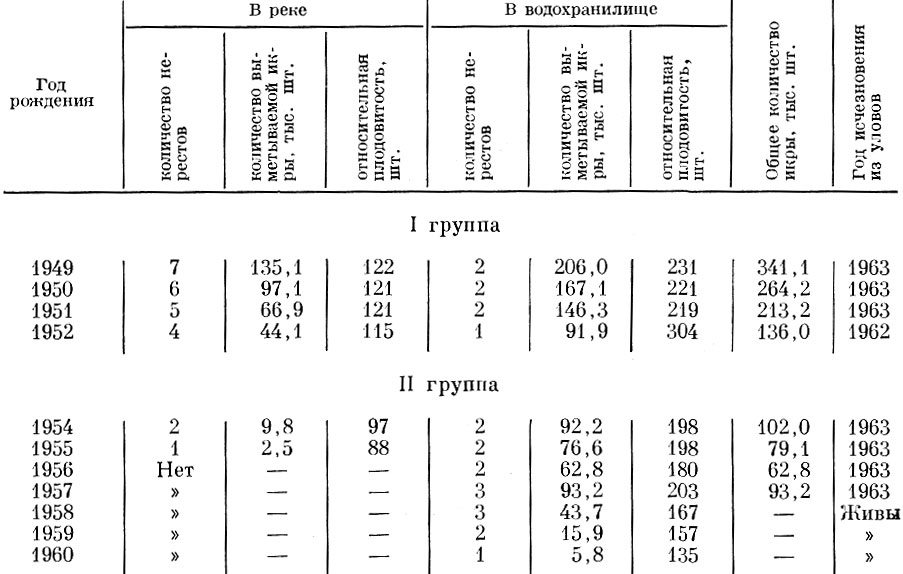
Таблица 31. Количество икры, выметываемое плотвой одного поколения, но живущей в разных условиях (по Спановской, Григораш и Лягиной, 1965)
Популяционная плодовитость плотвы в Можайском водохранилище возросла с 2,42 в 1960 г. до 6,25 тыс. икринок в 1961 г. Столь значительное увеличение плодовитости связано как с более ранним наступлением половой зрелости в связи с ускорением роста, так и с увеличением плодовитости одноразмерных рыб. Важно, что очень сильно возросла и относительная плодовитость у рыб одного и того же возраста и размера. Видимо, в дальнейшем по мере увеличения численности стада плотвы Можайского водохранилища плодовитость ее несколько снизится.
Сходная картина отмечена также и у синца Abramis ballerus (Pall.) Рыбинского водохранилища (Юровицкий, 1958).
Сильно изменилась плодовитость плотвы некоторых среднерусских водохранилищ за последние годы в связи с массовым развитием в них моллюска Dreissena, который стал существенным объектом питания рыб старших возрастов, что значительно повысило их обеспеченность пищей.
Это увеличение обеспеченности пищей плотвы крупных размеров (свыше 15 см длины) привело к значительному нарастанию ее плодовитости, причем это увеличение плодовитости почти незаметно на более мелких рыбах. Подобная картина отмечена в Учинском (Спановская, Григораш и Лягина, 1963) и Рыбинском водохранилищах (Володин, 1963) (рис. 14).
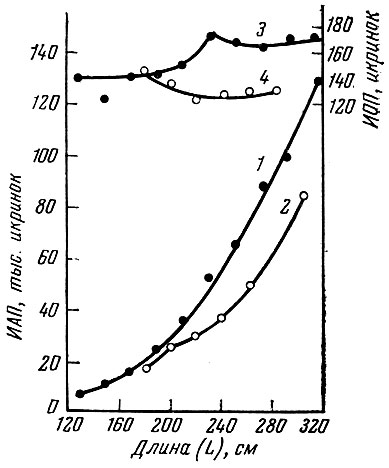
Рис. 14. Плодовитость рыбинской плотвы в 1961 и 1953 гг.: 1 - абсолютная плодовитость (ИАП) в 1961 г.; 2 - абсолютная плодовитость в 1953 г.; 3 - относительная плодовитость (ИОП) в 1961 г.; 4 - относительная плодовитость в 1953 г. (по Володину, 1963)
Плодовитость воблы Варваринского водохранилища выше, чем у воблы Мингечаурского водохранилища, что связано с ее лучшей обеспеченностью пищей и более высокой жирностью (Абдурахманов, 1971).
Изменения обеспеченности пищей популяций разных видов рыб в Куйбышевском водохранилище происходили неодинаково. Например, обеспеченность пищей плотвы, синца и густеры в 1960 г. (когда закладывалась плодовитость 1961 г.) улучшилась, а у леща она снизилась. Обеспеченность пищей рыб в Куйбышевском водохранилище определяется не только изменениями биомассы кормовой базы и численности кормящейся популяции. В значительной степени она меняется в связи с изменением термического и газового режима водоема (Гайниев, 1963). У леща Куйбышевского водохранилища происходит дальнейшее уменьшение плодовитости одноразмерных особей в связи с продолжающимся снижением обеспеченности пищей популяции этого вида (Цыплаков, 1969).
Б. К. Москаленко (1956) установил связь изменений плодовитости язя р. Оби с высотой уровня воды в реке. В годы, следующие за годом с высоким уровнем, плодовитость язя бывает более высокой, чем после годов с более низким уровнем (табл. 33). Эти изменения в плодовитости также, несомненно, связаны с изменениями обеспеченности пищей.

Таблица 33. Плодовитость язя р. Оби (Белогорье) (по Москаленко. 1956)
Существенно изменяется по годам плодовитость ряда видов карповых рыб и в Куйбышевском водохранилище (табл. 32).
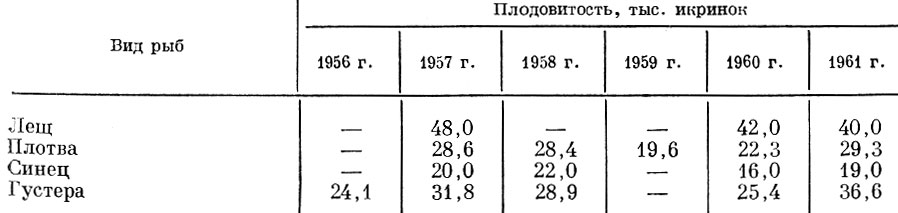
Таблица 32. Изменение плодовитости разных видов карповых рыб в возрасте 6+ в куйбышевском водохранилище за 1956-1961 гг. (по Гайняеву, 1963)
Сходное явление отмечено Н. П. Соколовым (1933) у северокаспийского сазана (табл. 34).

Таблица 34. Динамика плодовитости сазана (длиной 31-35 см) и продолжительность полойного периода в дельте волги (по Чугунову, 1928 и Соколову, 1933)
Однако связь плодовитости сазана и продолжительности полойного периода выражена четко не у всех размерных групп; у рыб старших возрастов она менее ясна.
После массовой гибели сазана в дельте Волги в результате эпизоотии (конец 50-х годов) численность его резко сократилась, что привело к улучшению обеспеченности пищей и значительному росту плодовитости. Так, если в 1923-1955 гг. средняя плодовитость самок сазана размером 41-45 см колебалась от 245 (1939 г.) до 321 тыс. икринок (1955 г.), то за период 1967-1968 гг. средняя плодовитость рыб этой же размерной группы составляла 429 тыс. икринок. Интересно, что за этот период у леща дельты Волги сколь-либо значительных изменений в плодовитости не: произошло (Васильченко, 1970).
Влияние изменения величины нагульных площадей и продолжительности сезона нагула на величину плодовитости отмечено О. А. Поповой (1965) также у щуки и окуня дельты Волги. В годы, благоприятные для нагула, абсолютная плодовитость оказывается выше. И у щуки, и у окуня повышение плодовитости связано, как и у многих других рыб, с повышением их упитанности и жирности (табл. 35).
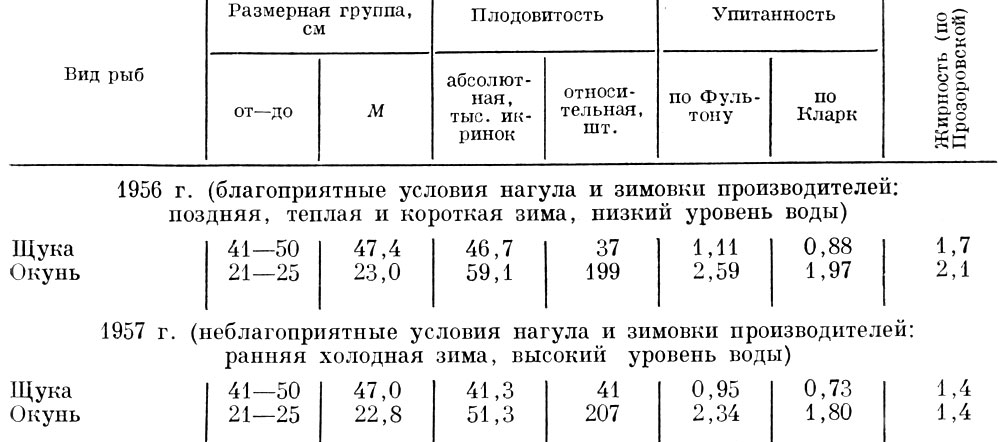
Таблица 35. Плодовитость, упитанность, жирность самок щуки и окуня в 1956 и 1957 гг. (нижняя зона дельты волги) (по Поповой, 1965)
У морской камбалы Platessa platessa (L.) Северного моря также удается подметить различия плодовитости в разные годы (Simpson, 1951). В 1948/49 г., например, плодовитость камбалы была несколько выше, чем в 1947/48 г. Симпсон указывает, что изменения плодовитости морской камбалы могут определяться как изменениями количества корма, приходящегося на одну особь, так и условиями питания, в первую очередь температурой.
В связи с изменениями условий жизни меняется как абсолютная, так и относительная плодовитость. Так, у многих рыб более мощные по численности возрастные и размерные группы из-за снижения их обеспеченности пищей оказываются и менее плодовитыми. Так, например, у обыкновенного гольца Nemachilus barbatulus (L.), по данным Либосварского (Libosvarsky, 1957), размерные группы, обладающие более высокой численностью, оказываются относительно и менее плодовитыми. Только у рыб длиной более 130 мм снижение как абсолютной, так и относительной плодовитости объясняется началом старения (табл. 36).
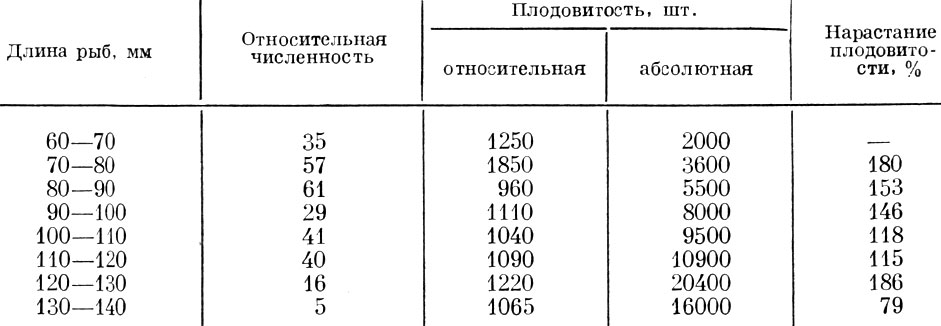
Таблица 36. Связь численности размерных групп и относительной плодовитости обыкновенного гольца Nemachilus Barbatulus (L.) чехословакии (по Либосварскому, 1957)
В тех случаях, когда обеспеченность популяции пищей меняется в связи с изменениями ее численности и у половозрелой части стада имеются расхождения спектра питания у рыб разного размера, как правило, снижается как абсолютная, так и относительная плодовитость рыб модальной размерной группы. Подобного рода примеры приведены выше. У разных видов рыб это отмечено также Г. Д. Поляковым (1962, 1968). Однако у тех рыб, спектр питания которых очень широк и разница между составом пищи половозрелых рыб разного размера выражена слабо, в годы высокой обеспеченности пищей плодовитость модальной группы почти не снижается. В годы же с низкой обеспеченностью пищей снижение плодовитости рыб модальной группы выражено сильнее. Это хорошо прослежено у беломорской наваги (Анухина, 1962, 1966; Поляков, 1968). У нее очень четко выражена обратная связь величины всей популяции и ее плодовитости, причем разница в плодовитости за отдельные годы очень велика (табл. 37).

Таблица 37. Средняя плодовитость наваги (по Анухиной, 1962)
Хуже изучены закономерности динамики плодовитости самцов. Известно, что как количество эякулята, так и концентрация спермиев в единице объема может очень сильно меняться. У отдельных видов рыб максимальная концентрация спермиев может превышать минимальную до 10 раз. Так же изменяется в больших пределах и объем эякулята (Турдаков, 1972). Г. Д. Поляков (1968), на основании анализа данных А. Ф. Турдакова для иссык-кульского чебачка, отмечает, что у самцов модальной по численности размерной группы концентрация спермиев ниже, чем у особей более малочисленных размерных групп.
Таким образом, анализ приведенного выше материала показывает, что в пределах одной популяции, как это правильно отмечают Ю. Е. Лапин и Ю. Г. Юровицкий (1959), действует зависимость: быстрый рост→раннее созревание→повышение плодовитости при повышении обеспеченности пищей и обратная картина при снижении обеспеченности пищей. В некоторых случаях повышение плодовитости может происходить и без ускорения роста, а быть связано с повышением жирности. Все эти изменения совершаются на фоне возрастных изменений, темп которых в свою очередь может приспособительно меняться в связи с изменениями условий жизни.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'