Возрастные изменения плодовитости
Изменения качества половых продуктов находятся в теснейшей связи с динамикой количественных характеристик воспроизводительных свойств популяции. Так же как и качество половых продуктов, плодовитость приспособительно меняется в пределах одной популяции. Она различна у разных популяций одного и того же вида и у разных видов, отражая условия их существования.
Важнейшая закономерность, которой подчиняются изменения плодовитости, это увеличение до определенных пределов абсолютной плодовитости по мере роста рыбы. Более крупные рыбы откладывают больше икры, чем мелкие, причем у большинства рыб корреляция плодовитости с массой рыбы выше, чем с длиной, а корреляция с длиной выше, чем с возрастом. Это видно на примере плотвы Рыбинского водохранилища (рис. 11).
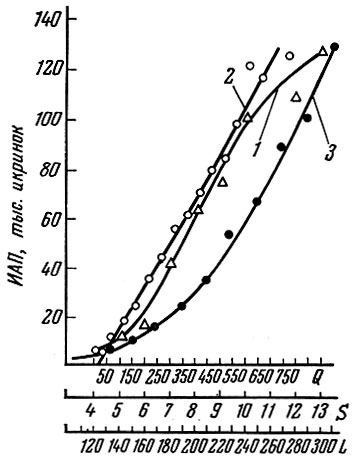
Рис. 11. Корреляция плодовитости плотвы с длиной тела (1), массой (2) и возрастом (3) рыб (по Володину, 1963)
Однако если увеличение плодовитости с ростом характерно для всех видов рыб, то характер этого увеличения, его пределы, когда повышение сменяется снижением плодовитости у старых рыб, оказываются специфическими для разных видов и отдельных популяций в пределах вида и отражают специфику взаимоотношений их со средой, специфику условий их жизни.
Как видно из данных табл. 18, плодовитость по мере роста рыбы, как правило, увеличивается все медленнее, а у старых рыб иногда обнаруживается даже снижение абсолютной плодовитости.
У рыб с длинным жизненным циклом (осетровые, сазан) плодовитость, как правило, быстрее нарастает у младших возрастов, затем нарастание несколько снижается и стабилизируется у средних размерных групп. Некоторые колебания в темпе нарастания плодовитости у средних размерных групп связаны с разным взаимным влиянием особей на обеспеченность рыб пищей в размерных группах разной численности. Нередко у более крупных рыб, численность которых мала, наблюдается некоторый подъем темпа нарастания плодовитости; это связано с улучшением обеспеченности пищей старших возрастов. Такой подъем обычно наблюдается в тех популяциях, в которых на увеличении плодовитости отрицательно сказывается взаимное влияние особей, и этот скачок в нарастании плодовитости приходится обычно на ту размерную группу, численность которой много меньше численности предыдущих размерных групп.
У самых крупных рыб плодовитость обычно резко падает. Однако характер нарастания плодовитости у разных видов весьма различен. У куринской севрюги (табл. 18) плодовитость быстро нарастает до того, как рыба достигает длины 140 см; после этого нарастание резко замедляется, а у рыб, достигших 190 см, плодовитость практически не увеличивается. Близкую картину удается наблюдать у аральского сазана, у которого максимальный темп нарастания плодовитости приходится на рыб длиной 45-50 см; затем темп нарастания плодовитости сильно снижается, а у рыб крупнее 60 см плодовитость даже несколько падает.
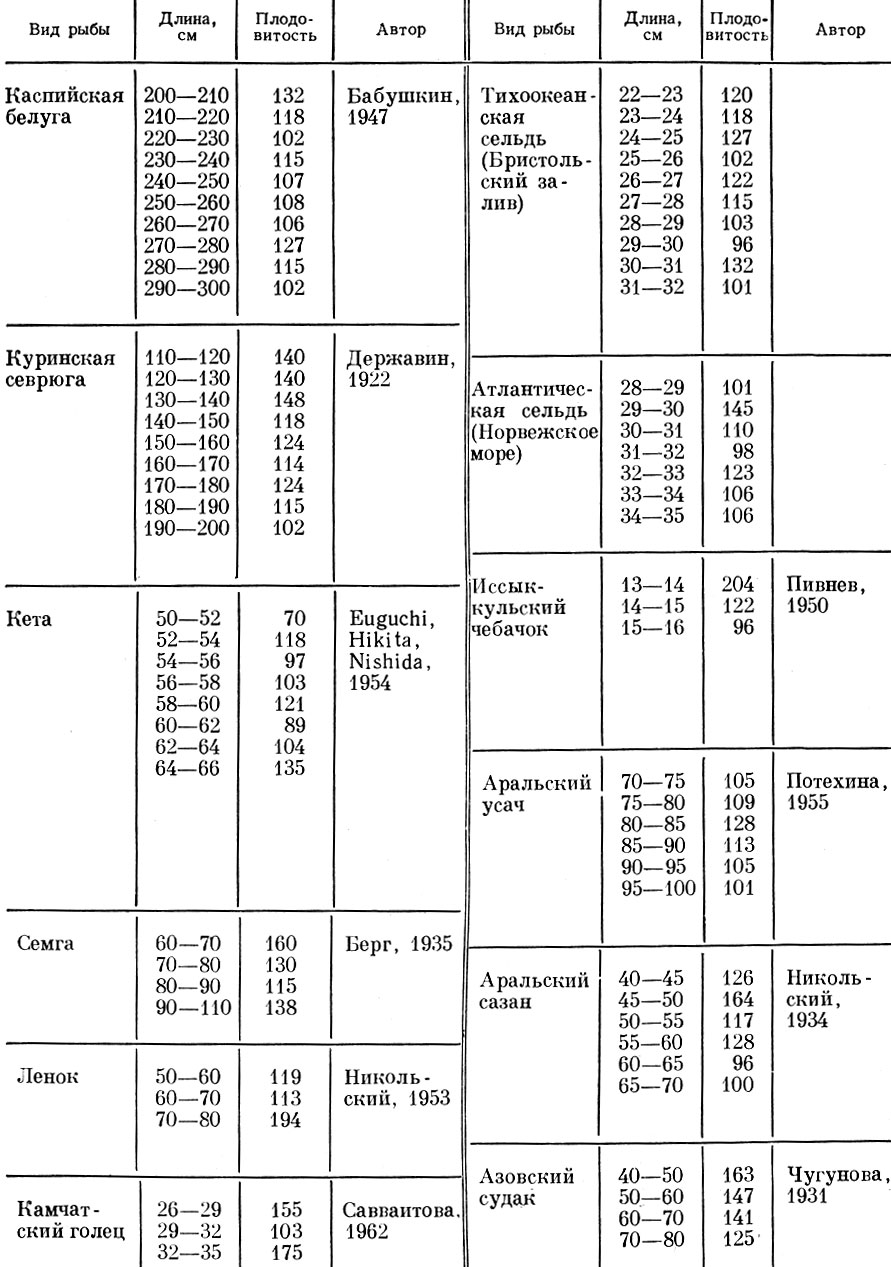
Таблица 18. Изменение абсолютной плодовитости разных видов рыб с увеличением их длины (в % к плодовитости предыдущей размерной группы)
У молодых и старых самок днепровского рыбца снижаются как общая плодовитость, так и число порций икры (Мороз, 1965).
Интересно, что у некоторых лососевых - семги, ленка - отмечается некоторый подъем темпа нарастания плодовитости у наиболее крупных рыб. У лососей, у которых нет повторного нереста (p. Oncorhynchus), закономерного снижения темпа нарастания плодовитости от более мелких к более крупным рыбам не наблюдается. Но у кеты, наоборот, нарастание плодовитости у наиболее крупных рыб усиливается.
У атлантического лосося плодовитость одноразмерных особей связана и с возрастом ската молоди из реки. Чем в более раннем возрасте особь переходит в покатную стадию, тем выше ее плодовитость (Евтюхова, 1965).
Совершенно естественно, что темп нарастания плодовитости, так же как и время наступления половой зрелости, меняется в связи с изменением обеспеченности пищей. Сравнение темпа нарастания плодовитости у сельди южной части Северного моря в 1933 и 1956 гг. обнаруживает весьма существенную разницу (табл. 19).
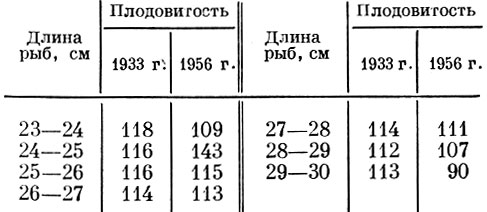
Таблица 19. Темп нарастания плодовитости сельди северного моря в разные годы (в % к плодовитости предыдущего размера)
В 1933 г. плодовитость сельди нарастала очень плавно и резкого снижения ее нарастания у рыб наиболее крупных размеров не наблюдалось. В 1956 г. наблюдались резкий скачок темпа нарастания плодовитости у мелких и значительно более быстрое падение у более крупных рыб. Интересно, что у самой крупной размерной группы сельди (29-30 см) и абсолютная плодовитость оказывается ниже, чем у предыдущей размерной группы. В 1933 г. подобного явления не наблюдалось.
У воблы Северного Каспия наблюдается сходная картина. В 1911 - 1912 гг. плодовитость нарастала медленнее, чем в 1946-1948 гг. (табл. 20), и максимум нарастания плодовитости в 1911-1912 гг., когда обеспеченность пищей популяции воблы была ниже, чем в 1946-1948 гг., приходился на рыб более крупного размера.

Таблица 20. Темп нарастания плодовитости северокаспийской воблы в разные годы, %
Как установили на примере плотвы Учинского водохранилища В. Д. Спановская, В. А. Григораш и Т. Н. Лягина (1963), улучшение обеспеченности пищей и связанное с этим изменение времени полового созревания и изменение темпа нарастания плодовитости в направлении перемещения максимума на более младшие возрастные группы обеспечивают разовое увеличение плодовитости популяции. В связи с тем что у рано созревающих рыб продолжительность жизни сокращается, при длительном изменении в сторону более раннего наступления половой зрелости популяционная плодовитость в ряде случаев может и снижаться по сравнению с плодовитостью популяций, состоящих из более поздно созревающих рыб. Очень четкие изменения в нарастании плодовитости отмечены у гольца Nemachilus barbatulus (L.) Чехословакии (см. табл. 36).
Таким образом, эта приспособительная реакция популяции на повышение обеспеченности пищей направлена обычно на кратковременное, иногда весьма значительное повышение воспроизводительной способности популяции, но она далеко не всегда может обеспечить длительное повышение плодовитости популяции.
Существенные возрастные изменения происходят и в величине относительной плодовитости. Относительная плодовитость, как правило, выше у впервые нерестующих особей, имеющих наиболее мелкую икру. Затем в течение более или менее длительного времени она остается относительно постоянной, иногда лишь несколько снижаясь у модальных по численности классов и затем более или менее резко снижается у старых особей. Ниже дано изменение относительной плодовитости у ленского осетра (по Л. И. Соколову, 1965).
| Длина, см | Плодовитость, тыс. икринок на 1 кг |
|---|---|
| 70-80 | 16,0 |
| 80-90 | 13,8 |
| 90-100 | 13,1 |
| 100-110 | 13,9 |
| 110-120 | 11,9 |
| 120-130 | 9,4 |
| 130-140 | 10,3 |
Сходный характер изменения относительной плодовитости с увеличением размеров и возраста отмечен и для многих других групп рыб: сельдевых (атлантическая сельдь, салака), лососевых (атлантический лосось, ленок), карповых (густера, лещ, сазан), окуневых (судак, окунь) и ряда др. Интересно отметить, что у некоторых видов рыб перед снижением относительной плодовитости у старых особей наблюдается ее некоторый подъем у предшествующих размерных или возрастных групп (Анохина, 1969; Поляков, 1968).
Возрастные изменения плодовитости связаны не только с количеством икринок, продуцируемых особями разного возраста, но и с изменением периодичности нереста. По-видимому, у многих видов рыб с длительным жизненным циклом старение связано не только со снижением относительной, а иногда и абсолютной плодовитости и увеличением количества резорбирующихся икринок, но и с изменением периодики нереста. Начало старения рыбы часто выражается в пропуске нерестового сезона. Так, у аральского леща, начиная с размеров 37-40 см, попадаются особи, которые нерестуют не каждый год (Бервальд, 1961). Аналогичное явление, видимо, наблюдается у старых особей других видов рыб - осетровых, сазана и некоторых других карповых, щуки, трески, судака и прочих. Естественно, что пропуск нереста у стареющих особей связан и с теми условиями, в которых живет популяция.
Возрастные изменения плодовитости самцов изучены еще плохо. Известно, что у большинства видов рыб с возрастом и размерами увеличивается количество продуцируемого эякулята. Для белого амура Ctenopharyngodon idella(Val.) отмечено повышение концентрацииспермиев в эякуляте у более крупных и старших по возрасту рыб (Попова, 1968). Однако эта закономерность свойственна не всем видам и, например, у иссык-кульского чебачка подобного явления не обнаружено (Турдаков, 1972).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'