Механизмы регуляции плодовитости
В формировании плодовитости рыбы четко выделяются два периода. Первый период - когда закладывается терминальный эпителий, т. е. на первом году жизни. В это время формируется, так сказать, общий уровень плодовитости особи, характерный для данной популяции (Simpson, 1951; Персов, 1963, и др.). Этот уровень плодовитости в дальнейшем подвергается значительным изменениям в связи с изменениями обеспеченности пищей. Условия нагула особи сильно отражаются на ее плодовитости и запасе желтка в икринке. Дальнейшая регуляция плодовитости связана с условиями нагула в предшествующий нересту нагульный сезон. Эти два периода формирования плодовитости хорошо выявляются на примере горбуши Oncorhynchus gorbuscha (Walb.), как акклиматизированной в Баренцевом и Белом морях, такие Южного Сахалина (рис. 4) (Персов, 1963).
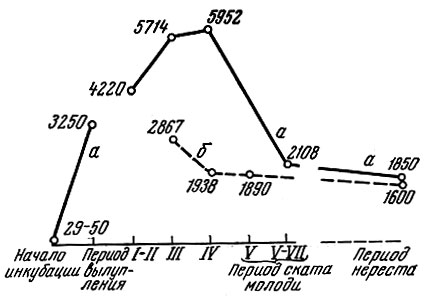
Рис. 4. Количество половых клеток у самок горбуши на разных этапах онтогенеза: а - Заполярье (1960 г.); б - Южный Сахалин (1959-1960 гг.) (по Персову, 1963)
Во второй период ее формирования плодовитость может изменяться как через регуляцию закладки новых овоцитов (Simpson, 1951), так и путем задержки или в случае повышения обеспеченности пищей ускорения перехода овоцитов фаз малого роста на стадию вакуолизации и наполнения желтком. Так, по В. Э. Беккеру (1957, 1958а), у обыкновенного карася при уплотненных посадках сокращается число зрелых овоцитов и выпадают одна или две порции нереста. Наполнение овоцитов желтком у рыб при низкой обеспеченности пищей происходит медленнее, чем при высокой.
Снижение плодовитости может осуществляться и путем атрезии овоцитов. Этот процесс может, видимо, происходить на всех стадиях их развития. Он стимулируется снижением обеспеченности пищей и имеет место как в экспериментальных условиях, так и у естественных популяций (Scott, 1962). Так, у Salmo gairdneri Richardson и в экспериментальных условиях при выращивании на голодном пайке, и в естественных уплотненных популяциях наблюдалось значительное снижение плодовитости вследствие резорбции части созревающих яиц. Ниже приведено изменение относительной плодовитости и рост овоцитов у Salvelinus fontinalis Mitch. Как показано В. Д. Владыковым (Vladykov, 1956), атрезия овоцитов и снижение плодовитости идут параллельно с ростом остающихся овоцитов.
| Средний диаметр икринки, мм | 1,62 | 2,52 | 3,50 | 4,25 |
| Относительная плодовитость | 3,94 | 3,17 | 2,73 | 2,51 |
| Развивающихся овотитов, % | 100 | 80,9 | 69,6 | 64,0 |
У мелкой осенней семги р. Варзуги плодовитость, определенная в устье реки, когда гонады находятся в III стадии, оказывается выше, чем определенная у этой же семги на IV стадии зрелости. У крупных, более упитанных особей осенней семги этого не наблюдается: у нее плодовитость, определенная у рыб с незрелыми гонадами и находящимися на IV стадии, оказалась почти одинаковой (Мельникова, 1964).
Количество резорбирующихся яиц у разных видов и в разных условиях, естественно, может быть весьма различным. Так, у Salvelinus fontinalis Mitch, процент резорбирующихся яиц может достигать 50, а у Еrуmyzon oblongus (Mitch.) даже 72 (Bagenal, 1967). Иногда, как это отмечено для салаки, атрезия части овоцитов наблюдается у особенно быстрорастущих или впервые созревающих особей (Анохина, 1969). Рассасывающиеся овоциты, возможно, выполняют определенную гормональную функцию. Так как у рыб старших возрастов большее количество поступающих в организм энергетических ресурсов обычно идет на образование жировых отложений, а у впервые нерестующих рыб основные ресурсы расходуются на белковый рост, то, естественно, что и голодание у рыб более старших возрастов (более жирных) в меньшей степени сказывается на плодовитости и качестве икры, чем у впервые нерестующих рыб.
Условия нагула сказываются и на воспроизводительной системе самцов. Причем у разных видов эта реакция может быть весьма различной. Так, у обыкновенного карася голодание в период сперматогенеза приводит к значительному снижению массы молок. Масса семенников снижается до десяти раз. У угря же преднерестовое голодание не сказывается на процессе сперматогенеза (Турдаков, 1972).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'