Показатели, используемые для оценки плодовитости
При анализе плодовитости рыб обычно пользуются несколькими показателями. Наиболее распространенным из них является индивидуальная, или абсолютная, плодовитость (ИП, или ИАП).
Под индивидуальной плодовитостью понимается количество находящихся в яичниках родительской особи икринок генерации данного года, т. е. тех икринок, которые должны быть выметаны в данном году. У рыб с единовременным икрометанием, как это правильно отмечает П. А. Дрягин (1949), в яичниках обычно имеются две группы овоцитов: крупные - генерации данного года, и мелкие - генерации последующего года или последующих лет. Наряду с этими двумя группами овоцитов часто имеются овоциты меньших размеров, чем овоциты первой группы; при благоприятных условиях жизни особи, мелкие овоциты догоняют в росте овоциты первой группы, а в неблагоприятных условиях могут резорбироваться или переходить в генерацию овоцитов следующего года.
В связи со сказанным число икринок в яичниках самки следует просчитывать на четвертой стадии развития гонад (по шестибалльной шкале), желательно непосредственно перед началом нереста.
Значительно сложнее определение индивидуальной плодовитости у порционно нерестующих рыб. У этих рыб в яичниках имеются овоциты разных порций созревания, и отличить овоциты последующих лет весьма трудно. Кроме того, как показала на примере барабули Mullus barbatus ponticus Essipov Л. С. Овен (1961), у многих порционно нерестующих рыб овоциты развиваются столь быстро, что даже просчет всех овоцитов, находящихся в зрелом яичнике, не отражает действительной индивидуальной плодовитости вида, которая оказывается сильно заниженной. Так, индивидуальная плодовитость барабули, определенная многими авторами путем непосредственного просчета овоцитов (Виноградов и Ткачева, 1950; А. И. Смирнов, 1949, и др.), оказалась много ниже числа фактически выметанных за сезон икринок. Определенная методом просчета плодовитость барабули оказалась равной 3631-87640 икринкам, а фактически за сезон было выметано около 1000000 икринок.
Столь быстрый темп развития икры свойствен главным образом порционно нерестующим рыбам тропиков и субтропиков. У порционно нерестующих рыб умеренных и высоких широт обычно выметываются две-три порции икры за нерестовый сезон. У этих видов методом простого просчета удается определить величину индивидуальной плодовитости.
У порционно нерестующих рыб, например у карпа, величину индивидуальной плодовитости удается с достаточной точностью определять по массе (весу) зрелых икринок, путем взвешивания самок до и после икрометания. Правда, при этом мы определяем так называемую рабочую плодовитость, т. е. количество выметанных икринок, но в большинстве случаев это количество оказывается очень близким к количеству икринок данной порции, находящихся в ястыках (Ирихимович и Зеленин, 1956).
Индивидуальная плодовитость связана как с размером гонад, так и с размерами икринок, т. е. в первую очередь с запасом питательных веществ в икринках.
Существенным показателем воспроизводительных свойств особей популяции вида является относительная плодовитость (ОП). Относительная плодовитость, т. е. количество икринок, приходящееся на единицу массы тела самки, отражает как состояние самки, так и качество продуцируемой ею икры. Показатель относительной плодовитости, как это правильно отмечал еще В. И. Мейснер (1933), позволяет вести сравнение особей и популяций внутри вида. Относительная плодовитость дает также представление о воспроизводительных свойствах данной особи, так как этот показатель уже не связан прямо с массой и длиной рыбы. Методически правильнее относить количество икры к массе тела без внутренностей, т. е. к массе порки, так как при вычислении относительной плодовитости по отношению к массе всего тела или массе тела без гонад сказывается искажающее влияние наполненности кишечника пищей, а в случае отнесения к общей массе - и массы гонад (Иоганзен и Загороднева, 1951).
А. И. Зотин (1961) попытался установить математическую связь между размерами икры и величиной относительной плодовитости. По его мнению, эта связь выражается следующим уравнением:
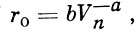 (1)
(1)где Vn - объем неоплодотворенного яйца;
а и b - константы, определяемые по приводимому в его статье графику соотношения размеров яиц и относительной плодовитости (ro).
А. И. Зотин находит, что, так как относительная плодовитость, определяемая как число икринок на единицу массы рыбы, меняется в пределах вида, то это понятие неудовлетворительно (почему?! - Г. Н.). Он предлагает другое и, как он сам пишет, несколько формальное определение относительной плодовитости как "коэффициент пропорциональности в линейной зависимости между абсолютной плодовитостью и весом рыбы".
А. И. Зотин считает, что у рыб есть три типа зависимости между абсолютной (г) и относительной (r0) плодовитостью, а именно:
1) r = r0Q (осетровые, сиги, некоторые карповые),
2) r = r0Q + С (обыкновенные и дальневосточные лососи),
3) r = r0Q - С (сарган, ставрида, губаны, бычки),
где:
Q - масса (вес) рыбы;
С - константа.
Сама по себе постановка вопроса о постоянстве относительной плодовитости неправильна. Плодовитость, в том числе и относительная, как и другие видовые свойства находится во взаимодействии со средой и может в известных пределах приспособительно меняться, отражая изменения условий жизни. Это относится как к относительной, так и абсолютной плодовитости. Мы согласны с теми авторами, которые считают, что состояние организма наиболее точно отражает показатель относительной плодовитости, выражаемый как число икринок на единицу массы тела рыбы. Ниже будет показано, что относительная плодовитость меняется как в онтогенезе, так и у разных популяций одного и того же вида; это имеет важное приспособительное значение.
Существенную ошибку допускает также А. И. Зотин, отрицая адаптивное значение абсолютной плодовитости и рассматривая ее только как прямую функцию изменений массы. Критика некоторых положений, высказанных А. И. Зотиным, и в первую очередь представления об инадаптивности индивидуальной плодовитости дана рядом авторов (Спановская, Григораш и Лягина, 1963; Персов, 1963; Поляков, 1968).
Несомненно, что увеличение плодовитости при увеличении массы рыбы в благоприятных условиях имеет важное приспособительное значение, направленное на увеличение численности стада. В то же время, как видно из излагаемого ниже, улучшение кормовых условий, вызывающее увеличение массы (веса) самок, далеко не во всех случаях влечет за собой увеличение плодовитости.
Несомненно, как абсолютная, так и относительная плодовитость, являющиеся видовыми признаками, имеют, как и другие видовые признаки, приспособительное значение.
Количество икры, которое производится особью, зависит не только от индивидуальной плодовитости, но и от числа икрометаний в течение жизни, т. е. от возраста полового созревания, продолжительности жизни или, точнее, продолжительности половой активности, и периодичности нереста.
С. А. Северцов (1941) предложил для оценки плодовитости животных формулу видовой плодовитости, в которой сделал попытку учесть вышеперечисленные моменты, определяющие плодовитость организмов. Формула С. А. Северцова имеет следующий вид:
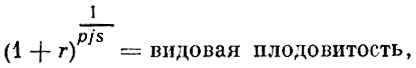 (2)
(2)где:
r - индивидуальная плодовитость;
р - период между двумя икрометаниями;
j - возраст наступления половой зрелости;
s - соотношение полов.
Определение плодовитости по формуле С. А. Северцова в известной степени отражает продукционную способность особей того или иного вида. Как показал С. А. Северцов на примере осетровых, их "видовая плодовитость" обратна индивидуальной плодовитости, т. е. рыбы с наименьшей индивидуальной плодовитостью обладают наибольшей видовой плодовитостью. Так, индивидуальная и видовая плодовитость осетровых, по С. А. Северцову (1941), следующая: у белуги индивидуальная плодовитость 664-4100000, показатель видовой плодовитости 0,070; у севрюги соответственно 35439-362935 и 1,502; у стерляди 16623-37632 и 4,286.
Формула С.А. Северцова вызывала ряд критических замечаний (Державин, 1947; Иоганзен, 1955; Поляков, 1971).
Так, Б. Г. Иоганзен указал, что, поскольку у рыб, как правило, соотношение полов близко 1 : 1, то этот показатель можно из формулы исключить. Формула, предложенная Б. Г. Иоганзеном, являющаяся видоизмененной формулой С. А. Северцова, имеет следующий вид:
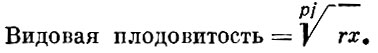 (3)
(3)В этой формуле приняты те же обозначения, что в формуле С. А. Северцова, только х - число икрометаний в течение жизни - введено Б. Г. Иоганзеном дополнительно.
Пользуясь своей формулой, Б. Г. Иоганзен расчислил видовую плодовитость ряда рыб Западной Сибири и на основе сравнения полученных величин пришел к выводу, что воспроизводительную способность разных видов можно объединить в три группы, различающиеся по величине этого показателя. Конечно, выделение этих трех групп в значительной степени условно и границы между ними выбраны произвольно.
Видовая, как и популяционная, плодовитость может приспособительно изменяться в определенных пределах из года в год. Неверно следующее утверждение Ф. И. Баранова (1929): "Так как естественное состояние водоема мыслится как состояние равновесия, то можно сказать: в естественных условиях видовая плодовитость всех рыб одинакова (независимо от числа выметываемых икринок) и равна 1". Ф. И. Баранов под видовой плодовитостью, видимо, понимал пополнение, что неправильно; кроме того, он, конечно, не прав, утверждая, что состояние водоема мыслится как равновесие. Известная динамическая уравновешенность кормности водоема и биомассы популяций рыб как раз и достигается регуляцией темпа воспроизводства, в том числе изменениями плодовитости популяции.
Критические замечания по поводу показателя видовой плодовитости были высказаны также В. С. Ивлевым (1953), который отмечал, что в формуле С. А. Северцова не учитывается возрастной состав стада, обычно несколько отличный в разных популяциях. B.C. Ивлев предложил свою формулу показателя популяционной плодовитости
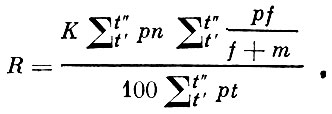 (4)
(4)где:
t - средний возраст;
t'- возраст наступления половой зрелости;
t"- возраст старости, т. е., по В. С. Ивлеву, времени прекращения размножения;
р - относительная численность данной возрастной группы (% от всех половозрелых рыб);
n - абсолютная плодовитость одной самки данного возраста;
f - число самок;
m - число самцов в популяции (средней пробе);
К - число икрометаний в течение года.
Пользуясь формулой, предложенной В. С. Ивлевым, Ю. И. Никаноров (1959) рассчитал популяционную плодовитость ряпушки ряда латвийских озер и показал, что иногда индивидуальная плодовитость недостаточно точно отражает действительную воспроизводительную способность популяции. Ю. И. Никаноров считает, что чем раньше наступает половая зрелость и чем больший процент составляют младшие половозрелые возрастные группы, тем выше популяционная плодовитость, тем выше воспроизводительная способность стада.
В формуле В. С. Ивлева, как и в формулах С. А. Северцова и Б. Г. Иоганзена, также не учитывается ряд моментов. В частности у многих рыб, как это, например, показано Э. А. Бервальдом (1961) в отношении леща, старение сначала выражается в изменении периодичности нереста, т. е. икрометание у старых особей происходит не каждый год, а с пропусками, обычно через год; этот период пониженной половой активности позднее сменяется ее утратой. Этот момент в формуле В. С. Ивлева не учитывается. Не учитывается В. С. Ивлевым и то, что у большинства рыб половая зрелость наступает не в одном возрасте, а часто возрастной ряд впервые нерестующих особей бывает сильно растянут. Формула В. С. Ивлева более пригодна в отношении рыб с коротким жизненным циклом, например ряпушки, чем в отношении рыб с длинным жизненным циклом и сложной структурой популяции.
Как показатель видовой плодовитости, введенный С. А. Северцовым (1941) и видоизмененный Б. Г. Иоганзеном (1955), так и показатель популяционной плодовитости, предложенный В. С. Ивлевым (1953), - это собственно не показатели плодовитости, а показатели, отражающие в той или иной степени темп воспроизводства популяции. Г. Д. Поляков (1971) для характеристики скорости воспроизводства стада рыбы предлагает пользоваться следующей формулой:
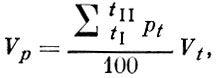
где:
Vp- популяционный показатель скорости воспроизводства (число самок на одну "исходную" самку в пересчете на один год);
Vt - средний индивидуальный показатель скорости воспроизводства самок, которые впервые нерестятся в возрасте t лет;
pt - относительная численность самок, которые впервые нерестятся в возрасте t лет (в % от всех впервые нерестующих самок);
tI - год первого нереста самок, созревших в наиболее раннем возрасте;
tII- год первого нереста самок, созревающих в наиболее старшем возрасте.
Под абсолютной популяционной плодовитостью, видимо, надо понимать общее число икринок, продуцируемое данной популяцией за нерестовый сезон. Эту величину определить трудно, так как для этого необходимо знать общую численность нерестовой популяции. Попытки подобного рода делались (Анохина, 1969), но, как правило, определение общей численности нерестовой популяции возможно лишь в первом приближении. В связи с этим чаще пользуются относительной популяционной плодовитостью (Никольский, 1960), которая выражается как средняя взвешенная плодовитость одной, ста, тысячи либо десяти тысяч рыб данной возрастной структуры нерестового стада. При этом количество рыб в каждой возрастной группе умножается на среднюю плодовитость рыб данного возраста; полученная путем подобного вычисления плодовитость рыб каждой возрастной группы суммируется с плодовитостью других возрастных групп; суммарные цифры и дают величину относительной популяционной плодовитости в том или ином году. Подобного рода расчеты были проведены мною для определения плодовитости сельди Северного моря за 1933 и 1956 гг. и воблы Северного Каспия за 1946 и 1947 гг. (Никольский, 1960). Как видно из приводимых ниже цифр, разница между плодовитостью одной и той же популяции оказывается весьма существенной. Так, в 1933 г. плодовитость ста сельдей при возрастной структуре стада этого года была 2100000 икринок, а в 1956 г. соответственно 2800000; у воблы Северного Каспия в 1946 г. - 4903900 икринок, а в 1947 г. - 2460500.
Таким образом, под относительной популяционной плодовитостью надо понимать среднюю абсолютную плодовитость самок разного возраста, длины или массы, вычисленную по относительной численности каждого возрастного размерного или весового класса. При этом, видимо, лучше всего пересчет вести на одну "среднюю" самку.
Для оценки плодовитости самцов обычно учитывается количество сперматозоидов в единице объема эякулята. Этот показатель, так же как и относительная плодовитость самок, позволяет сравнивать репродуктивную способность разных особей. Однако эта способность в очень большой мере зависит и от общего количества эякулята. Общее же количество продуцируемых самцами спермиев определить крайне трудно. Лишь в редких случаях удается определить общее количество выделяемого эякулята и для этого самцов приходится выдерживать в искусственных условиях, вследствие чего получаемые данные могут оказаться несколько искаженными.
А. Ф. Турдаковым (1968, 1972) предложена формула для определения общего объема спермы, продуцируемой самцом рыбы (опыты проведены на иссыккульской форели):
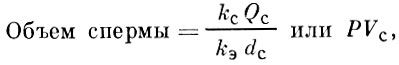
где:
kэ- концентрация спермиев в единице объема эякулята;
kс- концентрация спермиев в единице объема семенников;
Qc- масса семенников;
dc- удельный вес семенников;
Р - степень разбавления спермиев семенной плазмой при переходе из семенников в эякулят
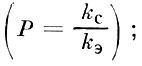
Vc - объем семенников.
Таковы те основные показатели, по которым мы можем судить о плодовитости рыб. Подробнее этот вопрос применительно к самкам рассмотрен Л. Е. Анохиной (1969).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'