Пищевые отношения внутри вида
Как известно, отношения между видами находятся в неразрывной связи с отношениями особей внутри вида. В процессе формирования вида вырабатывается определенная система внутривидовых связей, а также приспособлений, обеспечивающих регуляцию темпа воспроизводства в связи с меняющейся обеспеченностью пищей.
Основным приспособлением к расширению кормовой базы у вида является смена питания в процессе онтогенеза по мере перехода с одного этапа развития на другой. Смена состава пищи в онтогенезе имеет также значение и для обеспечения специфических морфоэнергетических процессов, протекающих в разные периоды жизни особи.
Уже разница в запасе желтка в икре и его изменчивость служат важным приспособлением, связанным с обеспеченностью пищей. У видов рыб с низкой обеспеченностью внешней пищей на ранних этапах онтогенеза запас желтка в икре больше, и на внешнее питание такие рыбы переходят при более крупных размерах, чем молодь рыб, приспособленных к высокой обеспеченности пищей. Это наиболее четко заметно при сравнении размеров желтка икры лососевых рыб, размножающихся в реках с низкой кормовой базой для молоди, и карповых, большинство которых размножается в стоячих и медленно текущих водоемах с более или менее богато развитым планктоном - основной кормовой базой молоди на ранних этапах развития. А среди карпоных у тех видов, которые размножаются в реках на течении, икра и свободные эмбрионы крупнее, чем у размножающихся в затишных участках озер и заводях рек, где развит планктон (Крыжановский, 1949).
В стоячих и медленно текущих водоемах отношение биомассы планктона к биомассе бентоса обычно составляет не менее 5:1. Если же сравнить годовую продукцию планктона и бентоса, то различие будет во много раз больше в пользу планктона. В то же время отношение биомассы особей рыб на этапах развития, приспособленных к питанию планктоном, к биомассе на этапах развития, питающихся бентосом, обычно оказывается близким к обратному. Так, у аральского леща отношение массы личинок в момент перехода на внешнее питание к массе родительской пары оказывается примерно 1 : 5,5. Надо иметь в виду, что при этом мы не учитываем биомассу тех неполовозрелых возрастов, которые питаются бентосом. Даже если учесть, что интенсивность потребления корма личинками леща много больше, чем взрослыми особями, то и тогда обеспеченность пищей особей на тех стадиях развития, когда они питаются планктоном, будет много выше,, чем у питающихся бентосом. Аналогичная картина наблюдается и у других рыб, питающихся бентосом во взрослом состоянии - воблы, белоглазки и др.
Сходное соотношение наблюдается и у морских рыб. Так, у трески и пикши оно оказалось примерно равным от 5 до 10 к 1 в пользу бентоснохищных стадий развития. Проведенный нами анализ спектра питания озерных рыб разных широт показал, что по мере роста особи спектр питания этих рыб становится все шире (Никольский, 1953). Так, например, у особей амурского чебака - Leuciscus waleckii Dyb. размером 5-10 мм, спектр питания состоит всего из двух групп компонентов, у особей 30- 40 мм - из восьми, а у взрослых - из 12. Такого же типа соотношение отмечено и у многих озерных рыб европейской части СССР, леща, плотвы и др. Естественно, однако, что широта спектра питания, находящаяся в тесной связи с обеспеченностью популяции данного размера или возраста пищей, связана и с численностью популяции рыб данного года рождения. Так, например, у чехони Рыбинского водохранилища, как показано А. Г. Поддубным (1955), спектр питания тех взрослых групп урожайных поколений, численность которых выше, оказывается шире, чем у более малочисленных возрастных групп.
Если у пресноводных озерных бентосоядных рыб спектр питания по мере роста становится все шире, то у тех пресноводных рыб, которые во взрослом состоянии перешли к питанию в эстуарных районах, наоборот, оставшаяся в пресной воде молодь оказывается хуже обеспеченной пищей, чем рыбы, перешедшие к нагулу в приустьевых пространствах. Так, у полупроходного леща Abramis hrama (L.) Северного Каспия спектр литания молоди в полоях состоит из двенадцати компонентов, а у взрослого леща на местах нагула в море - из шести. У воблы (и у молоди, и у взрослых рыб) спектр питания состоит примерно из одинакового числа компонентов.
У речных рыб, у которых все этапы развития проходят в реке с незначительным развитием планктонных организмов, обеспеченность пищей и взрослых рыб и молоди оказывается более или менее сходной, и спектр питания взрослых рыб и молоди по числу компонентов различается слабо.
У пресноводных рыб, в личиночном состоянии питающихся животной пищей, а во взрослом состоянии переходящих на питание растительностью или растительным детритом, обеспеченность пищей по мере роста рыбы увеличивается, а число компонентов в пище снижается. Это отмечено у ряда растительноядных и детритоядных рыб Амура - белого амура (Сысоева, 1953, Боруцкий, 1952), подуста-чернобрюшки и др.
У рыб водохранилищ обычно наблюдается картина, характерная для озерных бентофагов, т. е. спектр питания по мере роста рыбы расширяется, а обеспеченность пищей падает. Правда, при этом следует учитывать относительную численность возрастной группы: как это было отмечено применительно к чехони, группа с более высокой численностью сразу характеризуется расширением спектра питания и снижением обеспеченности пищей. Улучшение обеспеченности пищей у той или иной размерной или возрастной группы тотчас сказывается на сужении спектра ее питания. Так, например, если у плотвы в подмосковных естественных водоемах обычно спектр питания молоди уже, чем у взрослых рыб; то в водохранилищах, куда попала дрейссена, картина резко изменилась, и практически рыбы крупнее 16 см перешли на питание почти исключительно дрейссеной. Сходная картина наблюдается и у морских рыб. Так, например, если у молоди трески, держащейся в открытой части Баренцева моря, спектр питания уже, чем у более старших групп трески, то в губах, по данным Н. В. Мироновой (1956), у молоди трески и сайды наблюдается обратная картина, т. е. с возрастом у молоди этих рыб спектр питания становится более узким, причем это находится в прямой связи с численностью отдельных возрастных групп этих рыб в губах (табл. 2).
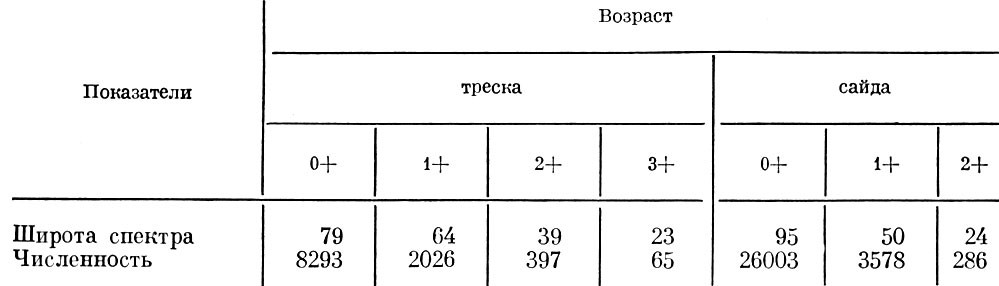
Таблица 2. Относительная численность и относительная широта спектра питания молоди трески и сайды разного возраста (по Мироновой, 1956)
Как видно, у старших возрастных групп, остающихся в губах в небольшом числе, спектр питания уже, чем у более младших. Особенно резко снижается численность и соответственно сжимается спектр питания у рыб в возрасте 2+, когда численность в губах по сравнению с предыдущей возрастной группой падает почти в 6 раз, а спектр питания сокращается почти в 2 раза. То же наблюдается и у сайды, когда численность рыб в возрасте 2+ в 10 с лишним раз меньше, чем рыб в возрасте 1+ , а спектр питания становится уже в 2 раза.
У личинок и молоди трески в открытой части Баренцева моря наблюдается связь накормленности и широты спектра питания. Чем выше индекс наполнения кишечника, тем уже спектр питания. У популяций молоди, хуже обеспеченной пищей, больше оказывается изменчивость их признаков и свойств (Никольский и др., 1968).
Расхождение мест нагула у рыб разного возраста тоже способствует повышению обеспеченности пищей популяции. Это имеет наибольшее значение для тех видов рыб, у которых в послеличиночный период развития возрастные изменения в составе пищи незначительны, и рыбы разного возраста питаются сходной пищей, например у атлантико-скандинавских сельдей, у беломорской наваги и др. Благодаря расхождению нагульных ареалов разные возрастные группы этих рыб выходят из противоречий из-за пищи.
Одним из приспособлений к повышению обеспеченности пищей у рыб является расхождение в составе пищи у особей разного пола. Это явление отмечено как у пресноводных, так и у морских рыб, причем разница в составе пищи не всегда связана с разницей в размерах самцов и самок. Часто одноразмерные самцы и самки питаются несколько различной пищей. Так, например, самки амурского чебачка Pseudorasbora parva Schl. в большем количестве, чем самцы, поедают личинок Glyptotendipes и Crycotopus gr. silvestris (Мухачева, 1950). Наблюдается существенная разница в составе пищи самцов и самок косатки-плети Liocassis ussuriensis Dyb.: у самцов в пище поденки значительно преобладают над ручейниками, у самок же их соотношение близко к 1 : 1 либо ручейников в пище больше (Егорова, 1958).
У леща в Азовском море самцы относительно в большем количестве, чем самки, поедают Nereis succinea, а самки - Hypaniola kovalevskii (Воробьев, 1937).
Разница между составом пищи самцов и самок наблюдается и у морских рыб. Кроме общеизвестного случая наличия паразитических самцов, питающихся соками тела самок у Ceratioidei, известны различия между составом пищи самцов и самок и у других морских рыб. Так, у беломорской трески Gadus morhua maris-albi Deriugin самцы относительно в большем количестве поедают ракообразных и червей. Различный характер питания самцов и самок беломорской трески также связан с некоторыми морфологическими различиями. Так, у самцов несколько короче голова, больше диаметр глаза, длиннее верхняя челюсть и брюшные плавники, больше высота хвостового стебля. Разница в составе пищи самцов и самок, связанная с морфологическими различиями, отмечена и у речной камбалы - Pleuronectes flesus L. (Микельсаар, 1958; Шатуновский, 1964).
Приведенные данные показывают, что у вида в процессе его становления вырабатывается ряд приспособлений, обеспечивающих расширение кормовой базы его популяций. Эти приспособления сводятся в первую очередь к расхождению спектра питания рыб разного размера и возраста. При этом возрастные группы, биомасса и численность которых бывает наибольшей, а обеспеченность пищей наименьшей, являются в большей степени эврифагами, т. е. имеют более широкий спектр питания. Широта спектра питания меняется с изменением обеспеченности пищей; это показано как наблюдениями в природе (Никольский, 1953), так и аквариальными опытами (Ивлев, 1955).
В процессе видообразования у видов вырабатывается ряд приспособлений к расширению кормовой базы при ухудшении обеспеченности пищей популяции. Основным из таких приспособлений является расширение изменчивости размеров и морфологической изменчивости одновозрастной группы рыб, что позволяет этой одновозрастной группировке осваивать более разнообразные корма. Возникающая в условиях пониженной обеспеченности пищей родительских особей разноразмерность икры связана с растянутостью выхода рыб из оболочки, а тем самым и растянутостью сроков начала внешнего питания (Никитинская, 1958). Разный запас желтка у рыб на этапе смешанного питания связан и с разной суточной ритмикой потребления корма. Как показала В. А. Григораш (1961) на примере личинок плотвы, особи с большим запасом желтка потребляют пищу с перерывами; у них есть перерывы в захватывании пищи в ночные часы. Личинки же с малым запасом желтка питаются круглые сутки.
Большая разноразмерность эмбрионов, выклюнувшихся из икры, менее обеспеченных пищей - менее упитанных - самок, при переходе этих эмбрионов на внешнее питание даже при одновременном выходе из оболочки обеспечивает начавшим питаться личинкам некоторое расширение их кормовой базы: более мелкие из них потребляют одни виды корма, а более крупные - другие.
Варьирование величины изменчивости размеров в одновозрастной популяции достигается не только увеличением или уменьшением разноразмерности икринок у самок, различно обеспеченных пищей. Изменение величины изменчивости размеров рыб в популяции связано и с изменениями обеспеченности пищей самой популяции. Как показано мною (Никольский, 1953) и Г. Д. Поляковым (1958, 1960, 1961, 1962), после того как ухудшение обеспеченности пищей достигнет определенной величины, среди особей популяции, росших до этого более или менее одинаково, начинает проявляться разница в скорости роста; часть особей начинает расти быстрее, а основная масса отстает. При этом резко проявляется асимметрия кривой распределения по размерам (рис. 1).
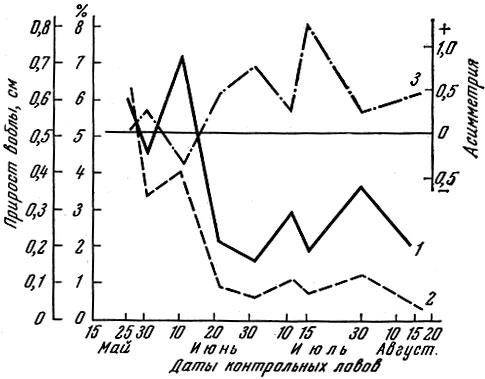
Рис. 1. Изменение скорости роста и величины асимметрии кривых размерного состава молоди воблы из рыбхоза Горелый, дельта Волги (по Полякову, 1960): 1 - абсолютный прирост за период между двумя контрольными ловами, мм за сутки; 2 - относительный прирост за тот же период, % среднего размера; 3 - величина асимметрии
Проявление в условиях пониженной обеспеченности пищей разнокачественное™ роста особей в популяции связано и с тем, что быстрорастущая часть особей раньше, чем медленно растущая, переходит на следующий этап развития. Способность популяции менять величину изменчивости размеров особей, ее слагающих, является важным приспособлением, обеспечивающим расширение спектра питания популяции, т. е. увеличение ее кормовой базы в условиях снижения обеспеченности пищей. В ряде случаев ухудшение обеспеченности пищей приводит к образованию карликовых форм. Так, например, у обыкновенного и серебряного карасей в озерах Северного Казахстана при снижении обеспеченности пищей популяция разделяется на более быстрорастущую и карликовую формы. Последняя растет примерно в 2 раза медленнее в длину и в 5-6 раз медленнее по массе. Пища карликовой формы менее разнообразна, чем быстрорастущей. В пищевом комке карликовой формы около 8 компонентов, а у более быстрорастущей - около 13. Основная пища карликовой формы - детрит, а более быстрорастущей - мотыль (Горюнова, 1962). При улучшении обеспеченности пищей карликовая форма исчезает. Возникновение карликовой формы в условиях ухудшения обеспеченности пищей наблюдается и у других рыб. Так, у глоссы Pleuronectes flesus luscus Pall, в Хаджибейском лимане, по-видимому, под влиянием низкой обеспеченности пищей в результате укороченного сезона нагула образовалась карликовая скороспелая форма (Замбриборщ, 1956).
У воблы в дельте Волги в условиях снижения обеспеченности пищей значительно увеличилась численность тугорослой скороспелой формы, отличающейся по характеру питания от более быстрорастущих рыб в популяции. Несомненно, что это изменение величины вариации размеров рыб в популяции осуществляется через перестройку обмена, но, к сожалению, более детально физиологический механизм этого явления не изучен. У нерки Oncorhynchus nerka (Walb.) в определенных условиях в озерах возникает мелкая жилая форма, которая достигает половозрелости в озере, не выходя в море. В популяции этой карликовой красной значительно преобладают самцы. Больше карликовых особей возникает от проходной формы нерки в годы с высокой обеспеченностью пищей в озере, когда рыба может достичь половозрелости, не выходя в море (Крохин, 1967).
Важным приспособлением к расширению кормовой базы является морфологическая разнокачественность признаков, связанных с питанием у особей в популяции.
В условиях высокой обеспеченности пищей, как это показано мною и В. А. Пикулевой (1958) на примере обыкновенного пескаря, вся популяция питается сходными кормами. В условиях же снижения обеспеченности пищей короткоусые особи пескаря в популяции переходят на один вид корма, а длинноусые на другой, вследствие чего спектр питания популяции в целом увеличивается и обеспеченность пищей повышается.
У популяций сига - Coregonus lavaretus (L.), живущих в условиях низкой обеспеченности пищей, особи с меньшим числом тычинок питаются одним видом корма, а более многотычинковые особи в основном другими кормами (рис. 2) (Решетников, 1964).
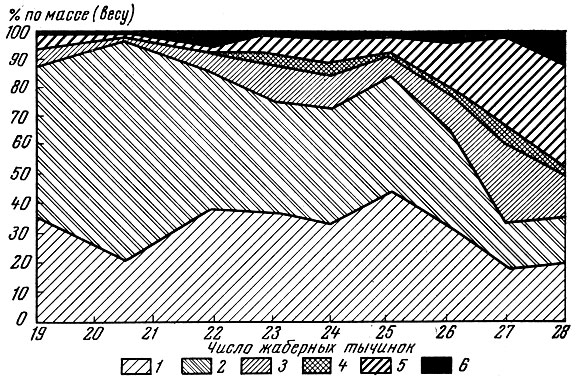
Рис. 2. Изменение характера питания сигов Чунозера в связи с числом жаберных тычинок (по Решетникову, 1964): 1 - моллюски; 2 - ручейники; 3 - двукрылые; 4 - воздушные насекомые; 5 - Cladocera; 6 - прочие организмы
У популяций, живущих в условиях низкой обеспеченности пищей, изменчивость признаков, связанных с добыванием и усвоением пищи, обычно выше, чем у популяций, хорошо обеспеченных пищей. Это отмечено мною при сравнении морфологических признаков аральского сазана, популяция которого хорошо обеспечена пищей, с признаками сазана из озер по Северному Узбою, где сазан живет в условиях очень низкой обеспеченности пищей.
Аналогичная картина наблюдается при сравнении живущих в разных условиях популяций обыкновенного карася. Популяция, живущая в условиях высокой обеспеченности пищей, обладает меньшей изменчивостью, чем популяция, живущая в условиях низкой обеспеченности пищей (Никольский, 1953). И в одной и той же популяции у поколений с высокой обеспеченностью пищей признаки и свойства менее изменчивы, чем у поколений, живущих в условиях низкой обеспеченности пищей. Так, например, у посаженной в один из прудов Звенигородской биологической станции Московского университета головешкиротана Percottus glehni Dyb. впервые годы (1953-1956) существования, пока ее численность не была высокой, изменчивость признаков и свойств в популяции была незначительной, спектр питания был узким и поедания собственной молоди не наблюдалось. По мере увеличения численности популяции изменчивость ее признаков увеличивалась, одновременно расширялся и спектр ее питания (табл. 3); в пище появилась собственная молодь (Спановская, Савваитова и Потапова, 1964).

Таблица 3. Изменение спектра питания и изменчивости головешки в разные годы
Важным приспособлением к повышению обеспеченности пищей являются нагульные миграции. При изменении плотности популяции меняется величина ее нагульного ареала (Марти, 1961). Нагульный ареал может, естественно, измениться и при стабильной численности, но если изменится кормность водоема или гидрологические условия на части нагульного ареала, то условия окажутся неблагоприятными для нагуливающихся рыб.
Сильнее всего "пульсация" нагульного ареала выражена у рыб, у которых есть значительные флюктуации (сардины, сельди, треска, пеламида). Флюктуации позволяют виду при наличии большого лабильного нагульного ареала и в случае значительного повышения обеспеченности пищей родительского стада через повышение урожайности потомства осваивать большой нагульный ареал (Никольский, 1974).
При сокращении по тем или иным причинам численности популяции иногда значительно сокращаются и ее нагульные миграции, и площадь нагульного ареала. Так, в последние годы уменьшается нагульный ареал в связи с сокращением численности у атлантико-скандинавских сельдей и у трески. Характерным примером сокращения площади нагульного ареала и протяженности миграций может служить тихоокеанская сардина, численность которой резко упала в результате неурожая 1939 г., и с 1941 г. резко уменьшились протяженность миграций и площадь нагульного ареала взрослых рыб.
Менее резко пульсация ареала выражена у тех рыб, у которых нет значительных флюктуации численности, например у камбал. Но и у них в годы высокой численности нагульный ареал расширяется, а в годы малой численности сокращается. При этом в зависимости от того, численность какой возрастной группы сократилась в связи с возрастными различиями в составе пищи и местах нагула, уменьшается та или иная часть ареала (Желтенкова, 1954).
Сходная картина наблюдается как у проходных, так и у пресноводных рыб.
У проходных рыб с обеспеченностью пищей в реках в значительной степени связаны начало, а часто и темп покатной миграции молоди и ее протяженность.
Очень важным приспособлением к повышению обеспеченности пищей популяции является стайный образ жизни в нагульный период, характерный для многих видов рыб (Никольский, 1955а). Особенно ярко стайный образ жизни выражен у планктоноядных, но часто он наблюдается и у донных рыб. В нагульный период стая обычно выполняет двоякую функцию: с одной стороны, непосредственно облегчающую отыскание и добывание корма, и с другой, оборонительную, а во многих случаях и миграционную. Все эти приспособительные функции стайного образа жизни связаны между собой (Мантейфель и Радаков, 1960). Непосредственное кормовое значение стаи заключается в том, что стая скорее, чем одиночная рыба, находит скопление кормовых организмов и легче сохраняет контакт с ним в случае, если скопление кормовых организмов подвижно. Одиночная рыба легче теряет скопление подвижных кормовых организмов, чем стая. Некоторые хищные рыбы образуют стаи и непосредственно для групповой охоты, что не только облегчает отыскание подвижной добычи и сохранение контакта с ней, но и непосредственно ее поимку. Групповая охота хищных рыб - довольно обычное в природе явление (Мантейфель и Радаков, 1960; Радаков, 1961, 1972).
В стае рыба, приспособленная к стайному образу жизни, обычно питается интенсивнее, чем в разреженном состоянии (Welty, 1934; Никольский и Кукушкин, 1943; Ивлев, 1955). Иногда на ранних стадиях развития рыбы, питающиеся планктоном, в стае питаются интенсивнее, чем в одиночку, а взрослые, питающиеся иными видами корма (бентос, детрит), в стае питаются менее интенсивно, чем в разреженном состоянии. Это, в частности, хорошо показал В. П. Воробьев (1937) на леще Азовского моря.
У рыб, образующих нагульную стаю, обычно питание и пищеварение совершаются со сходной ритмикой. Это позволяет всей стае одновременно начинать и одновременно заканчивать питание (Новикова, 1949, 1956), что облегчает рыбам в стае освоение скоплений кормовых организмов. Величина нагульной стаи имеет также не только оборонительное, но и непосредственное значение при питании. Величина стаи охотящихся в стае хищников непосредственно связана с величиной и поведением стаи их жертв и выработавшейся у данного вида хищников тактикой охоты (Мантейфель и Радаков, 1960). Размеры кормящейся стаи имеют значение при питании ташке у планктонофагов и бентофагов. Величина кормящейся стаи обычно соразмерна скоплению кормовых организмов; особенно четко это выражено у донных, кормящихся в стае рыб; у планктонофагов, кормящихся подвижным кормом, это тоже имеет место. У планктонофагов обычно в пределах кормящейся стаи рыбы еще группируются в небольшие косяки, состоящие из рыб, непосредственно взаимно ориентирующихся одна на другую.
Таким образом, объединение в стаи в нагульное время позволяет рыбе расходовать меньше энергии на поиск и добывание корма, т. е. повышает обеспеченность ее пищей.
Существенным приспособлением к повышению обеспеченности популяции пищей у некоторых видов рыб является питание себе подобными. Переход на питание особями того же вида имеет приспособительное значение-для повышения обеспеченности пищей в следующих трех направлениях:
1) когда крупные особи, питаясь своей молодью, осваивают через молодь такие корма, непосредственно питаться которыми крупные рыбы не приспособлены. Наиболее известным примером подобного рода могут служить обыкновенный окунь Perca fluviatilis (L.) и балхашский окунь Реrса schrenki Kessl., которые регулярно потребляют в пищу свою молодь. Это позволяет обыкновенному окуню часто жить в таких озерах, где других видов рыб нет и где бентос очень беден. Питаясь своей молодью, окунь через нее осваивает недоступный ему планктон;
2) когда молодь потребляет в пищу - непосредственно или через потребление беспозвоночных, питающихся трупами родительских особей, - тело своих родителей. Наиболее ярким примером в этом отношении служат дальневосточные проходные лососи, нерестующие в водоемах полугорного типа или олиготрофных озерах, где кормовая база для молоди очень бедна. Как показано Е. М. Крохиным (1959), в озерах Камчатки, где нерестует нерка Oncorhynchus nerka (Walb.), чем больше трупов нерки остается: после нереста в озерах, тем больше в воде фосфатов и тем богаче на следующий год в озерах развивается кормовой для молоди нерки планктон (рис. 3). В Амуре трупами производителей осенней кеты Oncorhynchus keta (Walb.) infrasp. autumnalis Berg, непосредственно питаются мальки кеты после выхода из нерестовых бугров до ската в море. В результате разложения трупов производителей горбуши Oncorhynchus gorbuscha (Walb.) и летней кеты Oncorhynchus keta typ. (Walb.) в нерестовых реках развивается бентос, которым питается молодь лососей. Таким образом, молодь проходных лососей, потребляя трупы родителей, расширяет свою кормовую базу. Отметим, что кормление на рыбоводных заводах молоди проходных дальневосточных лососей снёнкой - трупами отнерестовавших производителей - дает хорошие результаты. Близкое по характеру явление мы наблюдаем в озерах, где благодаря избыточно выпускаемой озерными рыбами сперме в местах нереста развивается более богатый планктон, служащий пищей выведшимся из икры личинкам (Le Cren, 1962);
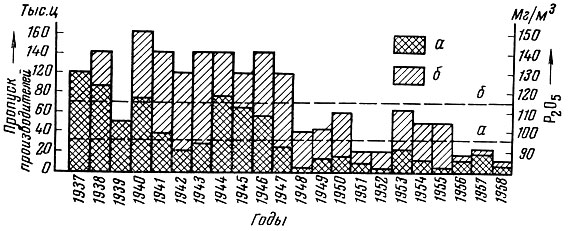
Рис.3. Пропуск красной в Дальнее озеро (а) и количество в нем фосфатов (б). Пунктиром показаны многолетние средние (по Крохину, 1959)
3) многие рыбы поедают собственную икру и молодь лишь в тех случаях, когда при высокой урожайности поколения старшие возрастные группы оказываются недостаточно обеспеченными пищей. Это явление довольно широко распространено среди рыб и является важным приспособлением для расширения кормовой базы популяции и регуляции ее численности в соответствии с кормовой базой водоема. Оно наблюдается у многих морских (треска, навага, Harpodon nehereus M. Clell, корюшка, скумбрия и многие др.) и пресноводных рыб (налим, щука, многие виды ушастых окуней - Centrarchidae и др.). По количеству своих икринок в кишечнике скумбрии Pneumatophorus japonicus Houtt. в известной степени можно судить о численности нерестовой популяции. Сводку данных о поедании рыбами особей своего вида см. у О. А. Ключаревой (1956).
Такое же биологическое значение имеют следующие явления: питание развивающихся эмбрионов соком тела родителей у живородящих акул, "молочное питание" плода у ската Pteroplatea, питание молоди слизью кожи родительских особей цихлиды - Symphysodon discus Heck. К этой же категории фактов относится и внутривидовой паразитизм у некоторых удильщиков подотряда Ceratioidei. Как известно, карликовые самцы удильщиков питаются соком тела самок. При этом кормовая база популяции не расширяется, но обеспечивается, при большой разреженности популяции этих рыб на глубинах, возможность нахождения друг друга особями разного пола (самцы прирастают ротовой полостью к телу самок).
Таковы приспособления, обеспечивающие популяции вида максимальное освоение кормовых ресурсов и изменение обеспеченности пищей в соответствии с меняющейся кормовой базой и численностью популяции.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'