Зообентос
Биологические особенности беломорского зообентоса удобнее всего рассмотреть на примере некоторых наиболее обычных представителей различных систематических групп беспозвоночных. В основу нашего очерка, как уже сказано, положены материалы личных наблюдений в Белом и Баренцевом морях в течение 1934-1940 и 1946-1953 гг.
Моллюски
Астаеa testudinalis (Müll.) по характеру своего географического распространения может считаться видом арктическо-бореальным (Дерюгин, 1915) или даже бореальным.
На Восточном Мурмане, а также в Воронке и в средней части Горла Белого моря этот вид широко распространен на каменистых грунтах нижнего отдела литорали и самого верхнего отдела сублиторали, предпочитая, однако, более или менее открытые участки берегов. Нередко он здесь живет и на литорали кутовых участков губ. Южнее Горла A. testudinalis встречается лишь в Бассейне, в Кандалакшском заливе и в северной половине Онежского залива; в остальных частях Белого моря он практически отсутствует.
Биологии A. testudinalis посвящена довольно обстоятельная статья Т. А. Матвеевой (1955а), материалами которой мы и воспользуемся.
При размножении A: testudinalis выметывает в воду зрелые половые продукты; оплодотворение и все последующие стадии развития эмбрионов и личинок протекают уже в водной толще, вне всякой связи с материнским организмом. На Восточном Мурмане, в Воронке и в северной части Горла более или менее зрелые продукты нередко имеются у самок в течение круглого года, но полной зрелости они достигают чаще всего только в июле. В годы с теплыми зимами созревание половых продуктов происходит уже в мае. Массовый вымет половых продуктов чаще всего начинается в июле и заканчивается в августе. В Бассейне и заливах Белого моря зрелые половые продукты в гонадах взрослых особей можно наблюдать, лишь в теплое время года. Размножение здесь протекает в более сжатые сроки и нередко оно бывает закончено уже в самых первых числах августа.
Более крупные размеры мурманских особей обусловлены, главным образом; значительным ускорением роста на первом году жизни: длина раковины в годовалом возрасте у беломорских особей почти во всех случаях меньшая, чем у мурманских. В последующие годы беломорские особи нередко растут с большей интенсивностью, чем мурманские, но отставание на первом году жизни оказывается решающим (табл. 39).
| Характер обитания | Возраст (годы) | Самая крупная особь |
Максимальный возраст |
|||||
| 1 | 2 | 3 | 4 | 5 | 6 | |||
| Открытые и полуоткрытые берега мелких островов Восточного Мурмана | ||||||||
| Длина раковины (в мм) | 5.4 | 10.5 | 15.0 | 20.5 | 24.5 | 26.3 | 33.5 | 10+ |
| Интенсивность роста (в %) | * | 94.4 | 42.9 | 36.7 | 19.5 | 7.8 | * | * |
| Открытые участки материкового берега | ||||||||
| Восточный Мурман Длина раковины (в мм) |
5.6 | 9.4 | 14.2 | 17.7 | 20.5 | 22.3 | 32.2 | 7+ |
| Интенсивность роста (в %) | * | 67.9 | 51.1 | 24.6 | 15.8 | 8.7 | * | * |
| Воронка Белого моря Длина раковины (в мм) |
4.2 | 7.3 | 10.7 | 13.3 | 16.2 | 18.2 | 25.5 | 6+ |
| Интенсивность роста (в %) | * | 75.2 | 46.6 | 24.3 | 21.8 | 12.3 | * | * |
| Северный берег Бассейна Длина раковины (в мм) |
4.3 | 7.3 | 10.7 | 13.3 | 16.2 | 18.2 | 20.7 | 7+ |
| Интенсивность роста (в %) | * | 69.8 | 46.6 | 24.3 | 21.8 | 12.3 | * | * |
| Полуоткрытые участки материкового берега | ||||||||
| Восточный Мурман Длина раковины (в мм) |
5.2 | 9.0 | 12.7 | 16.6 | 18.5 | 21.0 | 28.3 | 5+ |
| Интенсивность роста (в %) | * | 73.1 | 41.1 | 30.7 | 11.4 | 13.5 | * | * |
| Воронка Белого моря Длина раковины (в мм) |
4.7 | 8.1 | 11.9 | 15.1 | 18.0 | 20.5 | 25.5 | 8+ |
| Интенсивность роста (в %) | * | 72.3 | 46.9 | 26.9 | 19.2 | 13.9 | * | * |
| Северная часть Горла Длина раковины (в мм) |
4.8 | 7.7 | 12.3 | 16.4 | 20.4 | 22.4 | 25.5 | 7+ |
| Интенсивность роста (в %) | * | 60.4 | 59.7 | 25.0 | 24.4 | 9.7 | * | * |
| Западная часть Кандалакшского залива Длина раковины (в мм) |
4.3 | 7.5 | 10.4 | 13.6 | 16.8 | - | 19.4 | 5+ |
| Интенсивность роста (в %) | * | 74.4 | 38.7 | 30.8 | 23.5 | - | * | * |
| Восточная часть Карельского берега Длина раковины (в мм) |
3.9 | 6.4 | 8.5 | 10.4 | - | - | 21.6 | 3+ |
| Интенсивность роста (в %) | * | 64.1 | 32.8 | 22.4 | - | - | * | * |
| Закрытые участки губ | ||||||||
| Восточный Мурман Длина раковины (в мм) |
5.1 | 8.3 | 12.2 | 15.7 | 18.4 | 20.0 | 24.3 | 6+ |
| Интенсивность роста (в %) | * | 62.8 | 47.0 | 28.7 | 17.2 | 8.8 | * | * |
На Восточном: Мурмане максимальная продолжительность жизни, достигает 10+, а в Бассейне и заливах Белого моря - не более 7+. В наиболее угнетенных популяциях Восточного Мурмана особи живут до 5 +, а в Белом море только до 3+.
Littorina saxatilis (Olivi) служит одним из наиболее обычных и массовых представителей литоральной фауны моллюсков. Живорождение и ясно выраженные годовые кольца на раковине дают возможность детально и всесторонне изучить многие биологические закономерности, связанные с размножением, скоростью роста и продолжительностью жизни.
| Месяцы | Среднее количество зародышей на одну размножающуюся самку | ||||
| яйца и зародыши до сформирования велигер |
велигер | вполне сформировавшаяся молодь | всего | ||
| в яйцевых оболочках |
без яйцевых оболочек |
||||
| Восточный Мурман | |||||
| I | 6 | 4 | 10 | 40 | 60 |
| II | 9 | 9 | 30 | 55 | 103 |
| III | 0 | 0 | 0 | 71 | 71 |
| IV | 1 | 0 | 3 | 71 | 75 |
| V | 10 | 1 | 3 | 41 | 55 |
| VI | 51 | 9 | 1 | 1 | 62 |
| VII | 36 | 26 | 25 | 6 | 93 |
| VIII | 29 | 22 | 36 | 54 | 141 |
| IX | 6 | 8 | 21 | 91 | 126 |
| X | 8 | 7 | 11 | 44 | 70 |
| XI | 0 | 0 | 1 | 101 | 102 |
| XII | 1 | 1 | 2 | 35 | 39 |
| Белое море | |||||
| I | 1 | 2 | 4 | 52 | 59 |
| II | 1 | 3 | 2 | 54 | 60 |
| III | 1 | 1 | 5 | 49 | 56 |
| IV | 1 | 3 | 1 | 50 | 55 |
| V | 1 | 3 | 4 | 49 | 57 |
| VI | 0 | 2 | 4 | 63 | 69 |
| VII | 51 | 4 | 1 | 19 | 75 |
| VIII | 4 | 3 | 17 | 30 | 54 |
| IX | 5 | 7 | 21 | 38 | 71 |
| X | 0 | 8 | 16 | 46 | 70 |
| XI | 0 | 8 | 11 | 53 | 72 |
| XII | 0 | 3 | 8 | 54 | 65 |
На литорали Баренцева и Белого морей в выводковых сумках самок этого вида в любое время года имеются зародыши на той или иной Стадий развития (табл. 40-41). В теплое время года преобладают ранние стадии, а в холодное - поздние. Различие в цикле размножения мурманских и беломорских литторин состоит в следующем:
- Массовое рождение молоди на Мурмане происходит в мае-июне, а в Белом море только в июле.
- Период почти полного отсутствия в выводковых сумках сформировавшейся молоди на Мурмане. продолжается в течение 2 месяцев (июнь- июль), а в Белом море менее месяца (июль). Это обусловлено резко возросшей скоростью развития эмбрионов и сокращением продолжительности периода рождения молоди в Белом море. В результате этого накопление в выводковых сумках беломорских самок вполне сформировавшейся молоди начинается уже с середины лета и продолжается до следующей весны.
- Превращение всех или практически всех эмбрионов в вполне сформировавшуюся молодь на Мурмане завершается в феврале, а в Белом море уже в ноябре-декабре, но выражено здесь менее четко.
| Месяцы | Среднее количество зародышей на одну размножающуюся самку | ||||
| яйца и зародыши до сформирования велигер |
велигер | вполне сформировавшаяся молодь | всего | ||
| в яйцевых оболочках |
без яйцевых оболочек |
||||
| Восточный Мурман | |||||
| I | 0 | 0 | 0 | 63 | 63 |
| II | 0 | 0 | 0 | 71 | 71 |
| III | 0 | 0 | 8 | 72 | 80 |
| IV | 6 | 2 | 3 | 73 | 84 |
| V | 21 | 1 | 2 | 47 | 71 |
| VI | 47 | 21 | 6 | 14 | 88 |
| VII | 40 | 32 | 30 | 6 | 108 |
| VIII | 26 | 24 | 27 | 45 | 122 |
| IX | 17 | 10 | 16 | 47 | 90 |
| X | 10 | 4 | 14 | 56 | 84 |
| XI | 1 | 3 | 12 | 51 | 67 |
| XII | 0 | 0 | 6 | 60 | 66 |
| Белое море | |||||
| I | 0 | 3 | 6 | 51 | 60 |
| II | 0 | 2 | 4 | 53 | 59 |
| III | 0 | 2 | 3 | 45 | 50 |
| IV | 0 | 1 | 6 | 48 | 55 |
| V | 6 | 0 | 0 | 36 | 42 |
| VI | 20 | 11 | 12 | 8 | 51 |
| VII | 12 | 13 | 37 | 8 | 70 |
| VIII | 15 | 11 | 13 | 12 | 51 |
| IX | 1 | 3 | 11 | 61 | 76 |
| X | 1 | 2 | 8 | 90 | 101 |
| XI | 0 | 3 | 6 | 76 | 85 |
| XII | 0 | 3 | 7 | 73 | 83 |
Таким образом, основной особенностью цикла размножения беломорских литторин служит сокращение его наиболее активной части и завершение некоторых стадий в более сжатые сроки, чем на Мурмане (рис. 53), что особенно ясно выражено на закрытых участках побережья.
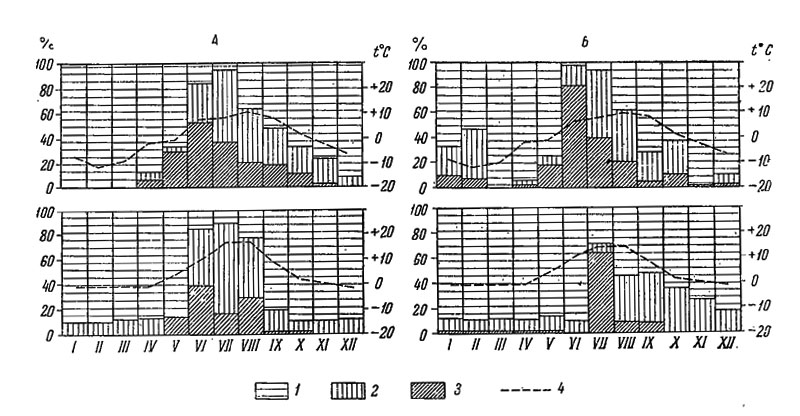
Рис. 53. Цикл размножения Littorina saxatilis на литорали Восточного Мурмана (вверху) и Карельского берега Белого моря (внизу). А - открытые и Б - закрытые участки литорали. 1 - яйца и зародыши до стадии велигер, 2 - велигер, 3 - вполне сформировавшаяся молодь, 4 - средняя месячная температура поверхности воды
Не меньшие различия обнаруживаются и при сравнении плодовитости мурманских и беломорских литторин (табл. 42). Совершенно очевидно, что беломорские литторины практически в любых местообитаниях размножаются при меньших размерах, чем мурманские, и имеют меньшую индивидуальную плодовитость. Относительная же плодовитость (количество зародышей на 1 г живого веса самки) беломорских литторин в общем более высокая, чем у мурманских. В среднем по всему изученному материалу на Мурмане она в конце лета составляет 414, а в Белом море 512. В наименее благоприятных условиях на Мурмане относительная плодовитость составляет 234 зародыша, а в Белом море 444.
| Характеристика местообитания | n | Диаметр раковины (в мм) | Индивидуальная плодовитость | Корреляция между диаметром домика и индивидуальной плодовитостью (r ± mr) | Количество зародышей на 1 г живого веса самки |
||||
| M±m | σ±mσ | υ±mυ | M±m | σ±mσ | υ±mυ | ||||
| Восточный Мурман Каменистые россыпи и скалы на открытых берегах мелких островов |
89 | 11.4 ± 0.1 | 1.3 ± 0.1 | 11.4 ± 0.09 | 110 ± 8 | 72 ± 5 | 65.5 ± 6.7 | +0.58 ± 0.07 | 234 |
| Каменистые россыпи и скалы на полуоткрытых берегах мелких островов | 204 | 13.5 ± 0.1 | 1.9 ± 0.1 | 14.1 ± 0.7 | 266 ± 10 | 140 ± 7 | 52.6 ± 3.2 | +0.75 ± 0.03 | 319 |
| Каменистые россыпи и скалы на открытых участках материкового берега | 99 | 8.5 ± 0.1 | 1.4 ± 0.1 | 16.5 ± 1.1 | 121 ± 5 | 53 ± 4 | 43.7 ± 3.6 | +0.62 ± 0.06 | 590 |
| Нижний отдел литорали на закрытых участках губ | 155 | 8.7 ± 0.2 | 2.9 ± 0.2 | 33.3 ± 2.1 | 115 ± 15 | 93 ± 1 | 89.3 ± 8.2 | +0.80 ± 0.03 | 477 |
| Верхний и средний отделы литорали на. открытых участках губ | 148 | 7.6 ± 0.1 | 1.7 ± 0.1 | 22.4 ± 1.4 | 67 ± 4 | 54 ± 3 | 80.6 ± 7.1 | +0.41 ± 0.07 | 462 |
| Белое море Каменистые россыпи и скалы на открытых участках мелких островов у Карельского берега и в северной части Онежского залива |
239 | 8.5 ± 0.1 | 1.8 ± 0.1 | 21.2 ± 1.0 | 110 ± 5 | 70 ± 3 | 63.6 ± 3.9 | +0.71 ± 0.03 | 537 |
| Каменистые россыпи на открытых и полуоткрытых берегах средней и южной частей Онежского залива | 281 | 8.5 ± 0.1 | 1.3 ± 0.1 | 15.3 ± 0.7 | 91 ± 3 | 47 ± 2 | 51.7 ± 2.7 | +0.49 ± 0.05 | 444 |
| Каменистые россыпи на открытых участках материкового берега и крупных островов в Бассейне и в Кандалакшском заливе | 242 | 6.8 ± 0.1 | 1.1 ± 0.05 | 16.2 ± 0.8 | 57 ± 2 | 34 ± 1 | 59.6 ± 3.5 | +0.56 ± 0.04 | 541 |
| Каменистые россыпи на закрытых участках губ Кандалакшского и северной части Онежского заливов | 194 | 7.5 ± 0.1 | 1.2 ± 0.1 | 16.0 ± 0.8 | 73 ± 3 | 44 ± 2 | 60.3 ± 4.0 | +0.59 ± 0.05 | 521 |
| Илисто-песчанистые пляжи на закрытых участках Поморского берега | 144 | 6.4 ± 0.1 | 1.2 ± 0.1 | 18.8 ± 1.1 | 46 ± 2 | 26 ± 1 | 56.5 ± 4.3 | +0.20 ± 0.07 | 515 |
Еще большие отличия беломорских самок от мурманских выявляются при сравнении размеров тела и плодовитости различных возрастных групп (табл. 43). На Восточном Мурмане самки впервые размножаются чаще всего в возрасте 2+ или даже 3 + , лишь в очень редких случаях в особо благоприятные годы единичные особи могут достигать половой зрелости в начале второго года жизни. За все время работ здесь было изучено много тысяч особей и только единственный раз была встречена самка, приступившая к размножению в конце первого лета жизни. В отличие от этого в Белом море самки обычно приступают к размножению в начале второго года жизни, а нередко и на первом году, т. е. в возрасте 0 + . Основную массу размножающихся самок на Мурмане составляют особи в возрасте от 3 + до 5 +, а в Белом море до 3 +. Относительная плодовитость беломорских самок во всех возрастных группах более высокая, чем у мурманских, особенно высокого уровня она достигает в молодом возрасте, но резко снижается к концу жизни. У мурманских самок снижение относительной плодовитости с возрастом выражено менее четко и не всегда.
| Возраст самок (годы) | Количество изученных размножающихся самок | Средний размер раковины (в мм)* | Индивидуальная плодовитость | Количество зародышей на 1 г живого веса самки |
|
| (в экз.) | в процентах | ||||
| Литораль открытых берегов мелких островов | |||||
| Восточный Мурман От 1+ до 3+ |
11 | 13.2 | 10.9 | 84 | 218 |
| От 4+ до 5+ | 58 | 70.0 | 11.4 | 102 | 224 |
| От 6+ до 7+ | 14 | 16.8 | 12.1 | 128 | 243 |
| Южная часть Бассейна Белого моря От 1+ до 3+ |
100 | 62.9 | 7.7 | 96 | 633 |
| От 4+ до 5+ | 48 | 30.2 | 8.8 | 113 | 497 |
| От 6+ до 7+ | 11 | 6.9 | 10.2 | 133 | 398 |
| Литоральные каменистые россыпи закрытых участков губ | |||||
| Восточный Мурман От 1+ до 2+ |
26 | 17.6 | 6.1 | 35 | 455 |
| От 3+ до 4+ | 104 | 70.2 | 6.8 | 59 | 537 |
| От 5+ до 6+ | 13 | 8.8 | 7.8 | 54 | 345 |
| Старше 6+ | 5 | 3.4 | 9.3 | 63 | 239 (?) |
| Карельский берег Белого моря От 1+ до 2+ |
34 | 36.6 | 6.4 | 53 | 604 |
| От 3+ до 4+ | 30 | 32.2 | 7.3 | 77 | 585 |
| От 5+ до 6+ | 25 | 26.9 | 8.7 | 80 | 368 |
| Старше 6+ | 4 | 4.3 | 8.0 | 81 | 486 (?) |
* Размером раковины у брюхоногих моллюсков и во всех других случаях считается диаметр последнего завитка (рис. 54).
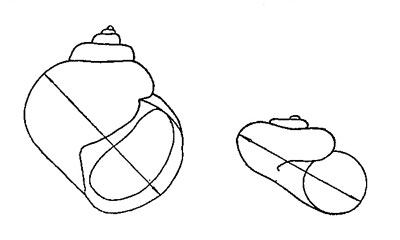
Рис. 54. Направления измерений раковины брюхоногих моллюсков (диаметр последнего завитка раковины)
Мурманские особи чаще всего обладают большей продолжительностью жизни и растут с большей скоростью, чем беломорские (табл. 44). Только на открытых берегах мелких островов беломорские особи получают некоторые преимущества перед мурманскими: здесь они имеют несколько большую длительность жизни и могут достигать более крупных размеров. Обращает на себя внимание то обстоятельство, что в первые годы жизни беломорские особи почти во всех случаях крупнее мурманских. С возрастом это преимущество беломорских особей постепенно исчезает, а затем превращается в свою противоположность, благодаря тому, что снижение скорости роста у них происходит раньше и более резко, чем у мурманских.
| Характеристика местообитаний |
Возраст (годы) | Самая крупная особь |
Максимальный возраст |
Интенсивность роста на 6 году/на 2 году |
|||||
| 1 | 2 | 3 | 4 | 5 | 6 | ||||
| Каменистые россыпи и скалы на открытых берегах мелких островов | |||||||||
| Восточный Мурман Диаметр раковины (в мм) |
4.4 | 6.7 | 8.4 | 9.4 | 10.4 | 11.2 | 13.7 | 6+ | 0.15 |
| Интенсивность роста (в %) | * | 52.3 | 25.4 | 11.9 | 10.6 | 7.7 | * | ||
| Белое море Диаметр раковины (в мм) |
4.3 | 6.6 | 7.5 | 8.7 | 9.4 | 10.1 | 14.8 | 7+ | 0.14 |
| Интенсивность роста (в %) | * | 53.5 | 13.6 | 16.0 | 8.0 | 7.2 | * | ||
| Каменистые россыпи и скалы на открытых участках материкового берега | |||||||||
| Восточный Мурман Диаметр раковины (в мм) |
2.5 | 4.0 | 5.3 | 6.1 | 7.2 | 7.8 | 13.0 | 18+ | 0.14 |
| Интенсивность роста (в %) | * | 60.0 | 32.5 | 15.1 | 18.0 | 8.3 | * | ||
| Белое море Диаметр раковины (в мм) |
3.3 | 4.8 | 5.7 | 6.2 | 6.7 | 6.8 | 10.3 | 8+ | 0.03 |
| Интенсивность роста (в %) | * | 45.5 | 18.7 | 8.8 | 8.0 | 1.5 | * | ||
| Каменистые россыпи и илисто-песчанистые пляжи на закрытых участках губ | |||||||||
| Восточный Мурман Диаметр раковины (в мм) |
3.0 | 4.6 | 6.2 | 7.2 | 8.0 | 8.5 | 12.0 | 11+ | 0.12 |
| Интенсивность роста (в %) | * | 53.3 | 37.0 | 16.1 | 11.1 | 6.2 | * | ||
| Белое море Диаметр раковины (в мм) |
4.4 | 6.7 | 7.8 | 8.4 | 8.8 | 9.4 | 10.5 | 7+ | 0.13 |
| Интенсивность роста (в %) | * | 52.3 | 16.4 | 7.7 | 4.8 | 6.8 | * | ||
Изложенное позволяет отметить следующие биологические особенности беломорских Littorina saxatilis, отличающие их от мурманских:
- Плотность поселений заметно понижается.
- Размножение остается также круглогодичным, но наиболее активная часть этого процесса (откладывание яиц, их развитие, рождение молоди) протекает в более сжатые сроки.
- Половая зрелость самок наступает в более раннем возрасте и при меньших размерах тела. В целом беломорские размножающиеся самки имеют меньшие размеры тела и меньшую индивидуальную плодовитость, но значительно более высокую относительную плодовитость.
- В большинстве местообитаний продолжительность жизни сокращается, уменьшается скорость роста и окончательные размеры тела.
- В первые годы жизни размеры тела почти во всех случаях более высокие, но снижение скорости роста с возрастом выражено значительно более резко, благодаря чему к концу жизни они становятся обычно мельче мурманских.
Littorina obtusata (L.) подобно предшествующему виду служит одним, из наиболее обычных и массовых представителей литоральных моллюсков. На неорганических субстратах литорали Восточного Мурмана плотность поселений может достигать 600 особей/м2 с биомассой 173.2 г; на сходных субстратах литорали Карельского берега Белого моря живет не более 120 особей/м2 с биомассой 9.0 г. На органических субстратах, т. е. на слоевище различных водорослей, плотность поселений на Восточном Мурмане составляет до 824; особей/м2 с биомассой 95.6 г, а у Карельского берега Белого моря 539 особей с биомассой лишь 24.0 г.
Подобное же уменьшение плотности поселений беломорских особей можно наблюдать и в отношении средних показателей для любого участка Бассейна или заливов.
При размножении L. obtusata делает кладку (рис. 55, 1), из которой по прошествии некоторого времени выходит вполне сформировавшаяся молодь. Кладка представляет собой студенистый, желатинообразный комочек с включенными в него зародышами (Baster, 1762, цит. по Lebour, 1937b; Pelseneer, 1911; Delsman, 1914; Hertling u. Ankel, 1927; Кузнецов, Матвеева, 1948). Кладки прикрепляются к поверхности слоевища водорослей или непосредственно к камням; на закрытых участках литорали кладки бывают обычно только на водорослях.
На литорали Восточного Мурмана первые кладки чаще всего появляются в самый разгар полярной зимы, в феврале (табл. 45); количество таких зимних кладок весьма ограничено, и заключенные в них эмбрионы почти не развиваются до июня. Период массового размножения занимает здесь время с мая по август, причем массовое появление новых кладок заканчивается в июле, а в течение августа и отчасти сентября завершается развитие эмбрионов в ранее отложенных кладках. Первая молодь выходит из кладок в июле и последняя - в августе, иногда в сентябре.
| Месяц | Всего кладок |
В том числе (в процентах) | Месяц | Всего кладок |
В том числе (в процентах) | ||||||
| яйца и эмбрионы до стадии велигер |
велигер | смешанные кладки (велигер и вполне сформировавшаяся молодь) |
вполне сформировавшаяся молодь |
яйца и эмбрионы до стадии велигер | велигер | смешанные кладки (велигер и вполне сформировавшаяся молодь) |
вполне сформировавшаяся молодь |
||||
| на Fucus vesiculosus | на Fucus serratus | ||||||||||
| I | 0 | 0 | 0 | 0 | 0 | I | 0 | 0 | 0 | 0 | 0 |
| II | 1 | 100 | 0 | 0 | 0 | II | 1 | 100 | 0 | 0 | 0 |
| III | 1 | 100 | 0 | 0 | 0 | III | 1 | 100 | 0 | 0 | 0 |
| IV | 1 | 100 | 0 | 0 | 0 | IV | 1 | 100 | 0 | 0 | 0 |
| V | 53 | 100 | 1 | 2 | 3 | V | 2 | 3 | 1 | 2 | 3 |
| VI | 267 | 92.9 | 5.5 | 1.6 | 0 | VI | 80 | 98.1 | 1.9 | 0 | 0 |
| VII | 155 | 24.9 | 19.3 | 31.2 | 24.6 | VII | 94 | 32.9 | 26.5 | 28.0 | 12.6 |
| VIII | 55 | 11.3 | 18.1 | 27.3 | 43.3 | VIII | 76 | 10.1 | 15.6 | 39.6 | 34.7 |
| IX | 1 | 0 | 0 | 0 | 100 | IX | 0 | 0 | 0 | 0 | 0 |
| X | 0 | 0 | 0 | 0 | 0 | X | 0 | 0 | 0 | 0 | 0 |
| XI | 0 | 0 | 0 | 0 | 0 | XI | 0 | 0 | 0 | 0 | 0 |
| XII | 0 | 0 | 0 | 0 | 0 | XII | 0 | 0 | 0 | 0 | 0 |
В отличие от этого, в Бассейне и заливах Белого моря сроки размножения этого вида сильно сокращаются: первые кладки появляются здесь лишь в середине июня и в подавляющем большинстве случаев исчезают уже в июле; лишь в наименее прогреваемых, открытых участках литорали Бассейна и северной части Онежского залива единичные кладки могут быть встречены еще и в августе. Вообще размножение у беломорских литторин происходит по существу при таких же высоких температурах, как и у живущих на литорали Западной Европы, в то время как на Мурмане они размножаются при самых низких температурах (табл. 46). Однако продолжительность периода икрометания с 7-9 месяцев в прибрежных водах Северного моря сокращается до 2 месяцев в Бассейне и заливах Белого моря. Основная масса кладок, как уже сказано, почти повсеместно откладывается на слоевище фукоидов; на 1 кг F. vesiculosus на литорали Карельского берега в первой половине июля насчитывается 28-52 кладки, тогда как на литорали Восточного Мурмана их бывает до 267 (табл. 45).
| Место наблюдений | Месяцы | Средние месячные температуры воды, при которых происходит икрометание |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Северный берег Бельгии | * | * | + | + | + | + | + | + | + | * | * | * | 7-17 | Pelseneer, 1911 |
| О. Гельголанд | * | + | + | + | + | + | + | + | + | + | * | * | 2-19 | Ankel, 1936 |
| Восточный берег Дании, Зунд | * | * | * | + | + | + | + | * | * | * | * | * | 4-18 | Thorson, 1946 |
| Восточный Мурман, район губы Дальне-Зеленецкой |
* | + | + | + | + | + | + | * | * | * | * | * | -0.2-8 | Наши наблюдения |
| Белое море, Бассейн и заливы | * | * | * | * | * | + | + | * | * | * | * | * | 11-16 | Наши наблюдения |
Примечание. + время икрометания.
Среднее количество зародышей в кладках Littorina obtusata широко изменчиво как в разное время года, так и в различных местообитаниях. Вот некоторые данные о среднем количестве зародышей в кладках этого вида:
- Различные местообитания закрытых участков литорали в губах Восточного Мурмана при отсутствии источников резких колебаний солености - от 66 до 154, а весной может подниматься даже до 354.
- Закрытые участки литорали в губах Восточного Мурмана при наличии источников регулярных и резких колебаний солености - не более 84.
- Различные местообитания на каменистых россыпях и скалах полуоткрытых участков литорали Воронки и Горла Белого моря (на Терском берегу) - 44 до 89, а весной увеличивается до 114-135.
- Различные местообитания на каменистых россыпях литорали северного берега Бассейна - от 51 до 72.
- Каменистые россыпи и скалы на открытых берегах островов северной части Онежского залива - 85.
- Каменистые россыпи и скалы на открытых берегах островов средней части Онежского залива - 37.
- Различные местообитания. Закрытые участки литорали в тубах Карельского берега - от 35 до 48.
Наблюдаемое в некоторых случаях заметное увеличение количества зародышей в весенних кладках обусловлено, очевидно, тем, что в это время года кладки делают преимущественно самки старших возрастных групп, индивидуальная плодовитость которых более высокая.
При содержании в садке (Кузнецов, 1947) самки в возрасте 5+ и старше за период с 29 апреля но 2 июня на Восточном Мурмане сделали в среднем по одной кладке каждая и за период с 2 июня по 28 июня еще по одной; после этого кладок больше не появлялось. Самки в возрасте от 3+ до 4+ единичные кладки сделали еще в мае (не более одной кладки на каждые три самки), за период с 28 июня по 26 июля они сделали в среднем еще по одной кладке и на этом их размножение закончилось. В первой половине августа некоторое количество кладок делают еще самки, впервые приступающие к размножению, кладки эти отличаются сравнительно мелкими размерами и малым числом заключенных в них эмбрионов. Таким образом, на Восточном Мурмане каждая самка в среднем делает от 1 до 2 кладок в течение года.
О фактических размерах индивидуальной плодовитости беломорских L. obtusata можно судить на основании следующего опыта: 28 июня (1947 г.) на литорали Карельского берега (в районе губы Гридиной) было собрано и помещено в два садка 20 копулирующих самок, по. 10 самок в каждом садке, вместе с ними помещались и самцы. Оба садка располагались в среднем горизонте литорали на закрытом участке губы. К 5 июля в одном садке самки сделали 11 кладок, а в другом 14, в дальнейшем кладок больше не появлялось, хотя самки содержались в садке вместе с. самцами до октября. Не было замечено и копуляции. Выход молоди из отложенных кладок начался 20 июля, а к 25 июля все кладки уже закончили свое развитие. Таким образом, в данном случае самки в среднем сделали по 1.25 кладки (около 50-60 зародышей), а развитие эмбрионов завершалось в течение 15-20 дней. Заметим, что наш опыт был начат в то время, когда всюду на литорали встречались кладки этого вида, следовательно весьма возможно, что собранные самки уже успели сделать некоторое количество кладок в естественных условиях.
В наших садках на Восточном Мурмане средний вес самки составлял 469 мг, а индивидуальная плодовитость 220 зародышей или 469 на 1 г живого веса. Беломорские самки имели средний вес 162 мг, индивидуальную плодовитость 62 и относительную плодовитость 384 зародыша. Сведения эти следует считать предварительными, характеризующими лишь порядок величин, а не различие между мурманскими и беломорскими особями. Небольшие наблюдения на открытых участках Карельского берега свидетельствуют о том, что там относительная плодовитость может подниматься до 680, т. е. становится значительно выше, чем на Мурмане.
Существенной биологической особенностью беломорских литторин служит также заметное сокращение периода размножения и ускорение эмбрионального развития (табл. 47).
| Стадия размножения | Побережье Англии, Франции и Германии | Проливы Балтийского моря | Восточный Мурман | Бассейн и заливы Белого моря |
| Созревание половых продуктов (месяцы) | III-VIII | III-VII | I-VII | VI-VII |
| Икрометание (месяцы) | III-IX | IV-VII | II-VII | VI-VII |
| Продолжительность развития эмбрионов (в сутках) | 21-28 | ? | 30-150 | 15-20 |
| Выход молоди из кладок (месяцы) | IV-X | ? | VII-IX | VII-VIII |
Уменьшение размеров тела беломорских особей обусловлено резким снижением скорости роста. Причем на первом году жизни они нередко бывают крупнее мурманских, но уже в возрасте 2 лет и старше становятся мельче их (табл. 48). Несмотря на несколько возросшую максимальную продолжительность жизни, беломорские особи во всех случаях остаются более мелкими по сравнению с мурманскими. Заметим, что более 90% особей как на Мурмане, так и в Белом море (из общего числа имеющих возраст от 2+ и старше) не живут более 5+ , поэтому указываемое увеличение продолжительности жизни беломорских особей практически не отражается на их численности.
| Характеристика местообитания |
Возраст (годы) | Самая крупная особь |
Максимальный возраст (годы) |
Интенсивность роста на 5 году/на 2 году |
||||
| 1 | 2 | 3 | 4 | 5 | ||||
| Каменистые россыпи и скалы на открытых берегах мелких островов | ||||||||
| Восточный Мурман Диаметр раковины (в мм ) |
4.9 | 8.2 | 10.8 | 12.3 | 13.2 | 15.8 | 7+ | 0.11 |
| Интенсивность роста (в %) | * | 67.3 | 31.7 | 13.9 | 7.3 | * | ||
| Белое море Диаметр раковины (в мм) |
5.3 | 7.1 | 9.0 | 9.4 | 10.2 | 13.0 | 8+ | 0.25 |
| Интенсивность роста (в %) | * | 34.0 | 26.8 | 4.4 | 8.5 | * | ||
| Каменистые россыпи и скалы на открытых участках материкового берега | ||||||||
| Восточный Мурман Диаметр раковины (в мм ) |
4.1 | 7.2 | 9.8 | 11.4 | 12.0 | 14.8 | 5+ | 0.07 |
| Интенсивность роста (в %) | * | 75.6 | 36.1 | 16.3 | 5.3 | * | ||
| Белое море Диаметр раковины (в мм) |
3.0 | 4.6 | 5.7 | 6.4 | 6.9 | 9.0 | 6+ | 0.15 |
| Интенсивность роста (в %) | * | 53.3 | 23.9 | 12.3 | 7.8 | * | ||
| Каменистые россыпи на закрытых участках губ | ||||||||
| Восточный Мурман Диаметр раковины (в мм ) |
4.5 | 6.7 | 8.4 | 10.1 | 10.7 | 14.8 | 7+ | 0.14 |
| Интенсивность роста (в %) | * | 40.9 | 25.4 | 20.2 | 5.9 | * | ||
| Белое море Диаметр раковины (в мм) |
4.6 | 6.6 | 8.0 | 8.5 | 9.0 | 10.4 | 9+ | 0.14 |
| Интенсивность роста (в %) | * | 43.5 | 21.2 | 6.2 | 5.9 | * | ||
Таким образом, биологические особенности беломорских L. obtusata, отличающие их от представителей этого же вида на Восточном Мурмане, состоят в следующем:
- Плотность поселений и биомасса на единицу площади заметно снижаются.
- Период размножения сокращается, а скорость эмбрионального развития увеличивается. Снижение численности и ускорение развития служат причиной общего уменьшения имеющихся на литорали кладок этого моллюска даже в самый разгар периода размножения.
- Индивидуальная плодовитость и размеры тела размножающихся самок заметно уменьшаются.
- Продолжительность жизни обычно несколько увеличивается, а средняя скорость роста значительно уменьшается, в результате чего беломорские особи всюду имеют более мелкие размеры. Скорость роста в первые годы жизни в некоторых случаях более высокая, но снижение ее с возрастом всегда более резкое.
Littorina littorea (L.) широко распространена на литорали побережий Англии, Франции, Германии, а вдоль побережий Норвегии поднимается до Восточного Мурмана и Белого моря. В прибрежных водах Восточного Мурмана и Белого моря этот вид не может быть отнесен к чисто литоральным обитателям, так как совершает ежегодные вертикальные миграции: летом он поселяется в нижних и средних отделах литорали, а с наступлением зимы уходит в сублитораль; впрочем на большую глубину никогда не опускается и зимует преимущественно у границы отлива.
Существенной особенностью популяции L. littorea на Восточном Мурмане служит практически полное отсутствие в ее составе особей моложе 5-6 лет. За весь многолетний период исследований и сборов здесь только один раз был найден экземпляр на третьем году жизни. Подобное же положение сохраняется в Воронке и Горле, вдоль Терского берега. В Онежском заливе, особенно в его южной части, особи на третьем году жизни и даже на втором (точнее говоря на второе лето жизни) не являются редкостью и иногда в массовом количестве встречаются на стеблях зостеры. Отсутствие молодых особей в популяциях, населяющих литораль Восточного Мурмана и части Белого моря, может свидетельствовать лишь о крайне медленном, далеко не ежегодном, появлении молоди или вообще об отсутствии способности к размножению. Наблюдения на Мурмане показали, что размножение, т. е. вымет самками вполне сформированных яйцевых капсул (рис. 55, 2) происходит ежегодно, следовательно, остается предположить, что отсутствие молоди обусловлено крайне слабым результатом размножения. Это, в свою очередь, может быть следствием отсутствия условий, необходимых для нормального развития эмбрионов и массовой гибели яйцевых капсул. Возможно, что такое положение наблюдается не всегда, а возникло лишь в последние годы и свидетельствует о временном угнетении L. littorea на литорали Восточного Мурмана и в некоторых участках Белого моря, являющихся краевой частью ее ареала. На литорали Восточного Мурмана численность этого вида весьма ограничена и на 1 м2 не бывает более 2-3 особей. В западной части Кандалакшского залива Белого моря на 1 м2 может встретиться уже 5-8 особей, причем площади, занятые поселениями L. littorea, здесь неизмеримо обширнее. В южной части Онежского залива плотность поселений увеличивается до 12-16 взрослых особей на 1 м2, а среди зарослей зостеры вдоль Поморского и Карельского берегов на 1 м2 нередко скапливается до 50-60 особей в возрасте от 1+ до 2+ и старше. Для сравнения укажем, что на литорали побережий Англии на 1 м2 обитает до 480 особей (Spoonera. Moor, 1940).
Икрометание у беломорских L. littorea происходит при таких же высоких температурах (6-15°), как и у берегов Англии, а на Восточном Мурмане - при самых низких температурах (5-10°), по сравнению со всеми другими участками географического ареала этого бореального вида (табл. 49). Вместе с этим, продолжительность периода икрометания с 8 месяцев у берегов Англии и Дании сокращается до 3 на Восточном Мурмане и до 2 месяцев на Карельском берегу Белого моря.
В Бассейне и заливах Белого моря более или менее массовые поселения L. littorea имеются (кроме Зимнего берега) почти повсеместно,, тогда как на Мурмане они обнаружены только на закрытых участках губ и заливов, например, в куту губы Ярнышной, в бухте Оскара и в губах Кольского залива (Гурьянова, Закс, Ушаков, 1928, 1930). На большом протяжении открытых берегов Восточного Мурмана и Воронки Белого моря этот вид практически отсутствует вовсе. Поэтому сравнение скорости роста и продолжительности жизни моллюсков в Баренцевом и Белом морях возможно только в отношении особей, населяющих закрытые участки губ и заливов (табл. 50). На Мурмане продолжительность жизни моллюсков достигает 28 лет, а диаметр раковины наиболее крупных из них - 33.4 мм (живой вес 9.85 г). Скорость роста и продолжительность жизни беломорских представителей этого вида в подобных же условиях значительно меньшие: самые крупные особи имеют раковину диаметром не более 27.5 мм (живой вес 6.06 г), а наиболее долговечные не живут более 16-17 лет. В сильно заиляемых участках Поморского берега L. littorea угнетена еще более: самые крупные особи имеют здесь раковину не более 18.4 мм при продолжительности жизни до 17 лет.
| Место наблюдений | Месяцы | Средние месячные температуры воды, при которых происходит икрометание |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Западный берег Англии, район Мильпорта | + | + | + | + | + | + | + | + | * | * | * | * | 6-15 | Tattersaal, 1920 |
| Южный берег Англии, район Плимута | * | + | + | + | + | * | * | * | * | * | * | * | 8-11 | Moor, 1937; Lebour, 1937 |
| О. Гельголанд | + | + | + | + | + | * | * | * | * | * | * | * | 2-12 | Ankel, 1936 |
| Побережье Бельгии | * | * | + | + | + | + | + | + | + | * | * | * | - | Lefevere, Leloup, Van Meel, 1956 |
| Восточный берег Дании, Зунд | + | + | + | + | + | + | + | + | * | * | * | * | 1-18 | Thorson, 1946 |
| Восточный Мурман, район губы Дальне-Зеленецкой |
* | * | * | * | * | + | + | + | * | * | * | * | 5-10 | Наши наблюдения |
| Белое море, район губы Гридиной | * | * | * | * | * | + | + | * | * | * | * | * | 6-15 | Наши наблюдения |
Примечание. + время икрометания.
В популяциях, населяющих наиболее открытые участки побережий Белого моря, продолжительность жизни L. littorea почти такая же, как на закрытых участках Восточного Мурмана, но скорость роста несколько снижена, в результате чего самые крупные особи имеют раковину диаметром 30.0-30.7 мм и живой вес от 7.43 до 7.85 г. В удаленных от Бассейна южных частях Онежского залива продолжительность жизни снижается до 17 лет, а размер раковины не превышает здесь 23.3 мм при живом весе 4.01 г. В большинстве популяций открытых участков беломорских берегов размеры раковины в конце первого года жизни значительно большие, чем у мурманских, а уменьшение размеров тела взрослых особей обусловлено резким замедлением их роста, начиная со второго года жизни. Если на Мурмане прирост раковины второго года жизни составляет 104.1-120.8%, то на открытых берегах Белого моря он колеблется в пределах от 63 до 80.9%. Интересно, что на закрытых участках губ, где скорость роста и продолжительность жизни в целом более низкие, прирост второго года жизни составляет до 111.4%. Иными словами, угнетение роста особей нередко наступает здесь позднее, чем в более благоприятных условиях на открытых берегах.
| Характеристика местообитания | Количество изученных особей |
Возраст (годы) | Самая крупная особь |
Максимальный возраст (годы) |
Интенсивность роста на 8 году/на 2 году |
|||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |||||
| Каменистые россыпи и илистро-песчанистые пляжи на закрытых участках губ и заливов при отсутствии источников регулярных и резких колебаний солености | ||||||||||||
| Западный Мурман Диаметр раковины (в мм ) |
28 | 4.8 | 10.0 | 14.1 | 16.8 | 19.5 | 21.0 | 21.7 | 23.2 | 27.5 | 13 | 0.07 |
| Интенсивность роста (в %) | * | - | 104.0 | 41.0 | 19.1 | 16.1 | 7.7 | 3.3 | 6.9 | - | ||
| Восточный Мурман Диаметр раковины (в мм ) |
20 | 4.8 | 10.6 | 16.5 | 20.2 | 22.2 | 24.4 | 26.2 | 27.0 | 33.4 | 28 | 0.03 |
| Интенсивность роста (в %) | * | - | 120.8 | 55.7 | 22.4 | 9.9 | 0.9 | 7.4 | 3.1 | - | ||
| Острова средней части Онежского залива Диаметр раковины (в мм) |
55 | 4.4 | 9.3 | 13.9 | 16.8 | 18.6 | 20.0 | 21.0 | 21.9 | 27.5 | 17 | 0.04 |
| Интенсивность роста (в %) | * | - | 111.4 | 49.5 | 20.6 | 10.7 | 7.6 | 5.0 | 4.3 | - | ||
| Губы Карельского берега Диаметр раковины (в мм ) |
71 | 5.5 | 11.1 | 15.3 | 17.9 | 19.3 | 20.1 | 20.9 | 21.7 | 27.2 | 16 | 0.10 |
| Интенсивность роста (в %) | * | - | 101.8 | 37.8 | 17.0 | 7.8 | 4.1 | 4.0 | 4.0 | - | ||
| Губы Поморского берега Диаметр раковины (в мм) |
499 | 4.5 | 7.2 | 9.7 | 12.0 | 12.1 | 13.0 | 13.9 | 14.7 | 18.4 | 17 | 0.10 |
| Интенсивность роста (в %) | * | * | 60.0 | 34.7 | 23.7 | 0.8 | 7.5 | 6.9 | 5.8 | - | ||
| Каменистые россыпи и илисто-песчанистые пляжи на открытых участках побережий | ||||||||||||
| Южный берег Бассейна Диаметр раковины (в мм ) |
95 | 6.2 | 10.6 | 13.9 | 16.5 | 18.5 | 19.8 | 20.9 | 21.8 | 30.7 | 22 | 0.06 |
| Интенсивность роста (в %) | * | * | 71.0 | 31.1 | 18.9 | 12.0 | 7.0 | 5.6 | 4.3 | - | ||
| Острова северной части Онежского залива Диаметр раковины (в мм) |
27 | 6.2 | 11.0 | 14.8 | 16.9 | 18.2 | 19.6 | 20.7 | 21.4 | 30.0 | 27 | 0.05 |
| Интенсивность роста (в %) | * | * | 77.4 | 34.5 | 14.2 | 7.7 | 7.7 | 5.6 | 3.4 | - | ||
| Лямицкий берег Диаметр раковины (в мм ) |
146 | 4.7 | 8.5 | 12.1 | 14.6 | 16.8 | 18.9 | 19.6 | 20.5 | 27.9 | 18 | 0.06 |
| Интенсивность роста (в %) | * | * | 80.9 | 42.4 | 20.7 | 15.1 | 13.7 | 3.8 | 4.8 | - | ||
| Острова южной части Онежского залива Диаметр раковины (в мм) |
192 | 5.4 | 8.8 | 11.2 | 12.8 | 14.1 | 15.0 | 15.3 | 16.5 | 23.3 | 17 | 0.12 |
| Интенсивность роста (в %) | * | * | 63.0 | 27.3 | 14.3 | 10.2 | 6.4 | 2.0 | 7.8 | - | ||
Основываясь на приведенных материалах, можно определить некоторые биологические особенности беломорских L. littorea:
- Численность, плотность поселений и биомасса значительно увеличиваются. Весьма характерно присутствие особей всех возрастных групп, тогда как на Мурмане молодь практически не встречается.
- Период икрометания в Белом море сокращается до 2 месяцев против 3 на Восточном Мурмане и 7-8 у берегов Западной Европы.
- В Белом море молодь появляется ежегодно, а на Мурмане, по-видимому, крайне редко, хотя вымет половых продуктов и развивающиеся яйцевые капсулы в планктоне наблюдаются там ежегодно.
- На закрытых участках Белого моря скорость роста, длительность жизни и размеры тела значительно меньшие, чем на Мурмане. На открытых участках Белого моря скорость роста, длительность жизни и размеры тела близки к тому, что наблюдается на Мурмане.
- На первом году жизни размеры тела беломорских особей крупнее, чем у мурманских. Снижение скорости роста с возрастом у беломорских особей выражено более резко, и поэтому к концу жизни они всегда более мелкие, чем мурманские.
Lacuna pallidula Da Costa в наших водах является одним из типичных представителей бореальной фауны. У южных берегов Англии (Plymouth Marine Fauna, 1931), а также у берегов Франции она живет исключительно или преимущественно на литорали (Gallien et Larambergue, 1936). Лишь в Орезунде (Thorson, 1946) она встречается, по-видимому, чаще всего в сублиторали. Вдоль побережий Восточного Мурмана, западного берега Воронки и Горла Белого моря массовые поселения этого вида на литорали встречаются только летом, причем нередко они занимают в одинаковой мере и сублитораль до глубины 1-3 м. В течение зимы L. pallidula на литорали Восточного Мурмана обычно отсутствует; в это время года она живет преимущественно в сублиторали на глубинах до 10 м. Такое сезонное изменение характера местообитания обусловлено изменением многих факторов внешней среды, из которых укажем сильные осенние и зимние штормы, снегопады и низкие зимние температуры воздуха (во время отлива), заставляющие моллюска покинуть литораль и на всю зиму перейти в сублитораль, где колебания условий жизни в значительной мере сглажены. Заметим, что яйцевые кладки этого вида имеются на литорали в любое время года и способны безболезненно переносить воздействие этих факторов, губительных для взрослых особей (Полянский, 1950, 1955).
В Бассейне и заливах Белого моря характер сезонных изменений в распределении L. pallidula меняется. Толстый ледяной припай надежно укрывает литораль и ее обитателей от губительного воздействия низких зимних температур воздуха и сильных штормов, и это дает возможность беспрепятственного существования здесь L. pallidula в течение всей зимы. С началом весны частые и резкие колебания солености, резкие суточные колебания температуры воды и воздуха заставляют моллюска опускаться в сублитораль, где он и живет на слоевище фукоидов, образующих нередко мощные заросли у границы отлива. Осенью и зимой значительная часть зарослей макрофитов разрушается, и в верхнем отделе сублиторали многих участков Бассейна и заливов Белого моря скапливается много гниющих растительных остатков, вызывающих более или менее ясно выраженный дефицит кислорода. По-видимому, это обстоятельство и заставляет L. pallidula уходить из сублиторали и подниматься на сравнительно хорошо аэрируемую литораль, которая в это время года отличается удивительной стабильностью в отношении многих факторов внешней среды.
На литорали открытых участков побережий Бассейна, где скопления растительных остатков почти не бывает, а колебания условий жизни летом в значительной мере ослаблены, этот вид в течение круглого года живет в самом нижнем отделе литорали и сезонных миграций не совершает. По нашим наблюдениям, в Двинском заливе и в южной поло вине Онежского залива L. palliadula не встречается.
Кладка L. pallidula (рис. 55, 3) представляет собой прозрачный студенистый комочек линзообразной формы, диаметром от 2 до 7 мм.
Количество зародышей в кладках изменяется в различных местообитаниях:
| * | Среднее количество зародышей в одной кладке |
| Восточный Мурман | |
| Литораль открытых участков | 48-59 |
| Литораль закрытых участков | 33-55 |
| Белое море | |
| Каменистые россыпи и скалы на литорали северной части Горла |
40 |
| Каменистые россыпи и скалы на открытых берегах мелких островов северной части Онежского залива |
41 |
| Каменистые россыпи на литорали полуоткрытых участков Карельского берега |
39 |
| Каменистые россыпи на литорали закрытых участков Карельского берега |
23 |
Кладки мурманских особей в среднем содержат от 33 до 59 зародышей; более крупные кладки свойственны литорали открытых участков побережья. В Белом море каждая кладка в среднем содержит от 23 до 41 зародыша, причем и здесь количество зародышей в кладках уменьшается по мере удаления от открытого моря.
| Месяцы | Всего | В том числе (в процентах) | ||||
| с яйцами | все стадии, предшествующие велигер |
велигер | смешанные кладки, преимущественно велигер и вполне сформировавшаяся молодь |
вполне сформировавшаяся молодь |
||
| Восточный Мурман, закрытые участки губ | ||||||
| I | 34 | 44.6 | 55.4 | 0 | 0 | 0 |
| II | 85 | 30.8 | 40.9 | 28.3 | 0 | 0 |
| III | 27 | 29.3 | 60.7 | 10.0 | 0 | 0 |
| IV | 91 | 15.7 | 34.7 | 49.6 | 0 | 0 |
| V | 63 | 13.5 | 39.3 | 46.2 | 1.0 | 0 |
| VI | 68 | 14.9 | 46.0 | 31.2 | 7.9 | 0 |
| VII | 66 | 13.0 | 38.5 | 23.9 | 19.1 | 6.5 |
| VIII | 83 | 4.7 | 22.2 | 28.3 | 29.6 | 15.2 |
| IX | 26 | 0 | 11.1 | 17.5 | 46.7 | 24.9 |
| X | 13 | 0 | 10.0 | 45.0 | 30.0 | 15.0 |
| XI | 6 | 0 | 0 | 25.0 | 50.0 | 25.0 |
| XII | 140 | 100 | 0 | 0 | 0 | 0 |
| Зимнее состояние кладок (I-III) | ||||||
| Восточный Мурман | 34.9 | 42.0 | 27.4 | 13.5 | 3.2 | |
| Белое море | 0 | 25.4 | 71.2 | 0 | 3.4 | |
| Летнее состояние кладок (VI-VIII) | ||||||
| Восточный Мурман | 13.9 | 42.0 | 27.4 | 13.5 | 3.2 | |
| Белое море | 14.3 | 76.2 | 9.5 | 0 | 0 | |
На слоевище многих литоральных макрофитов Восточного Мурмана, главным образом фукоидов и ламинарий, кладки L. pallidula в значительном количестве могут быть легко обнаружены в любое время года.
В качестве примера рассмотрим сезонные изменения в количестве кладок на слоевище Fucus inflatus (табл. 51). Свежеотложенные кладки появляются здесь с декабря по август, т. е. период икрометания у самок этого вида продолжается непрерывно в течение почти 9 месяцев. Развитие эмбрионов в кладках протекает крайне медленно, и выход молоди начинается не ранее конца июня; последняя молодь покидает кладки в ноябре. В других местообитаниях с небольшими отклонениями сохраняется в общих чертах такой же цикл размножения (Кузнецов, 1948). В среднем можно принять, что на Восточном Мурмане икрометание протекает с декабря по август, а выход молоди - с июля по ноябрь. Продолжительность развития эмбрионов в кладках, отложенных в декабре, составляет около 200, а отложенных в июле-августе около 70-90 суток.
В Воронке и Горле Белого моря цикл размножения очевидно остается таким же, как на Восточном Мурмане.
В восточной части Карельского берега Белого моря и на островах северной части Онежского залива из 59 кладок, собранных с конца января до середины марта, в 15 эмбрионы были на стадии дробления, в 42 на стадии велигер, а в двух находилась уже вполне сформировавшаяся молодь. Различие в зимнем состоянии мурманских и беломорских кладок становится отчетливым при сравнении процентного соотношения между кладками с разными стадиями развития эмбрионов (табл. 51).
Из этого сравнения следует, что в отличие от Мурмана в Белом море зимой новых кладок не появляется, развитие эмбрионов в ранее отложенных кладках (ноябре-декабре) заканчивается в течение января- марта и отчасти в апреле (?).
С началом весны в Белом море начинается второй период размножения, продолжающийся с июня по август. Кладки, собранные в северной части Онежского залива в течение второй половины июня и первой половины июля, в отношении стадии развития заключенных в них эмбрионов оказались отстающими по сравнению с кладками на Восточном Мурмане (табл. 51).
Выход молоди из летних беломорских кладок происходит в конце июля - в августе.
Приведенный материал показывает следующие различия в цикле размножения L. pallidula в центральных частях Белого моря и на Восточном Мурмане:
| * | Восточный Мурман | Белое море |
| Икрометание | XII-VIII | XI-XII и V-VII |
| Выход молоди | VII-XI | I-II (IV?) и VIII |
Иными словами, непрерывный процесс размножения с декабря по ноябрь на Восточном Мурмане в центральной части Белого моря распадается на два весьма коротких: с ноября по март (апрель?) и с июня по август.
На Восточном Мурмане развитие эмбрионов протекает при средних месячных температурах воды не более 10°, в то время, как на побережье Западной Европы и в Белом море средние месячные температуры воды поднимаются до 14-18°. На Восточном Мурмане кладки L. pallidula встречаются в течение круглого года, а в Белом море и на побережье Западной Европы - лишь часть года: в Белом море они отсутствуют осенью, с сентября по октябрь, а в районе острова Гельголанда - летом и осенью, т. е. с июля по ноябрь (табл. 52).
| Место наблюдений | Месяцы | Средние месячные температуры воды, при которых протекает развитие |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| О. Гельголанд | + | + | + | + | + | + | * | * | * | * | * | + | 2-14 | Hertling и Ankel, 1927; Ankel, 1936 |
| Восточный берег Дании, Зунд | * | * | * | + | + | + | + | * | * | * | * | * | 6-18 | Thorson, 1946 |
| Восточный Мурман, район губы Дальне-Зеленецкой |
+ | + | + | + | + | + | + | + | + | + | + | + | -0.2-10 | Наши наблюдения |
| Белое море, район губы Гридиной | + | + | + | + | + | + | + | + | * | * | + | + | -1.5-16 | Наши наблюдения |
Примечание. + время икрометания и нахождения кладок.
Жизненный цикл L. pallidula на побережье Восточного Мурмана завершается в течение одного года (Кузнецов, 1948); лишь отдельные особи могут иметь продолжительность жизни до 18-20 месяцев. Половая зрелость наступает в возрасте 4-12 месяцев, и после первого же размножения особи обычно погибают. Каждая самка в течение всей своей жизни делает одну, редко две кладки.
На закрытых участках Бассейна и заливов Белого моря жизненный цикл сокращается: молодь, вышедшая из кладок летом, к началу зимы достигает половой зрелости, размножается и погибает. В течение второй половины зимы и начала весны L. pallidula в таких местообитаниях бывает чаще всего представлена лишь кладками. Половая зрелость наступает здесь в возрасте 5-6 месяцев, а продолжительность жизни не превышает 8-9 месяцев.
Резко изменяется положение на открытых участках побережий Белого моря. Молодь, вышедшая из кладок в течение лета, к зиме достигает половой зрелости и размножается. После размножения, в отличие от того, что наблюдалось на Восточном Мурмане и на закрытых участках беломорских берегов, особи не погибают, а продолжают жить и размножаться еще в течение 12-18 месяцев. Таким образом, продолжительность жизни достигает в этих условиях 2-2.5 лет (и даже 3 + ), а самки участвуют в размножении не менее 4 раз. Вот, например, сведения о возрастном составе такой популяции (в %):
| Возраст (годы) |
Зимой | Летом |
| 0+ | 92.5 | 58.3 |
| 1+ | 7.5 | 38.9 |
| 2+ | - | 2.8 |
Как видно, более 40% особей имеют продолжительность жизни более одного года.
К концу первого года жизни беломорские особи имеют диаметр раковины в среднем 3.0, к концу второго лета 4.5 и на третье 6.7 мм. Обращает на себя внимание тот факт, что возрастная элиминация идет преимущественно за счет быстро росших особей: в конце первого лета, жизни молодь имеет диаметр раковины от 1.4 до 6.8 мм, а на втором году жизни сохраняются лишь те, которые имели размер от 1.7 до 2.6 мм. Самые крупные особи в Белом море обычно имеют диаметр раковины не более 7.5 мм, но изредка могут быть встречены особи с диаметром раковины до 9.2 мм и в возрасте 3+.
На Восточном Мурмане, несмотря на сокращение продолжительности жизни, моллюски достигают более крупных размеров. Например, на закрытых участках губ самые крупные особи могут иметь раковину диаметром до 9.0 мм. Интересно, что несмотря на это, скорость роста особей на Восточном Мурмане и в Белом море, по-видимому, практически одинакова. Если на открытых участках побережий Бассейна в конце первого лета жизни особи достигали размера 6.7 мм, то на Восточном Мурмане в этом же возрасте диаметр раковины не превышал 7.0 мм.
Различие же между двумя популяциями состоит в том, что в Белом море, как уже сказано, возрастная элиминация идет преимущественно за счет быстро росших особей; на Восточном Мурмане столь резкой разницы в выживаемости быстро и медленно росших особей не наблюдалось.
В общих чертах биологические особенности беломорских представителей L. pallidula состоят в следующем:
- Численность и плотность поселений в Белом море значительно сокращаются, а в некоторых районах поселения этого вида отсутствуют вовсе.
- Период икрометания сокращается в общем с 9 месяцев на Восточном Мурмане до 4 месяцев. Выход молоди из кладок на Мурмане продолжается в течение 5 месяцев, а в Белом море - из летних кладок в течение 1, а из зимних - 4 месяцев.
- Количество зародышей в кладках иногда такое же, как на Восточном Мурмане, но в большинстве случаев меньшее.
- На закрытых участках побережий длительность жизни особей в Белом море и на Восточном Мурмане приблизительно одинакова, но на открытых и полуоткрытых участках беломорские особи - могут жить в, 3-4 раза дольше мурманских. Скорость роста беломорских особей замедлена, но благодаря увеличению длительности жизни они иногда могут достигать таких же и даже более крупных размеров.
Lacuna vincta (Montagu) в пределах своего ареала, ограниченного преимущественно бореальной областью, занимает лишь узкую прибрежную полосу и чаще всего служит представителем литоральной фауны.
На восточном Мурмане, в Воронке и в северной части Горла Белого моря L. vincta живет также среди водорослей нижнего отдела литорали, но количественно всегда преобладает среди ламинарий и некоторых других макрофитов ниже линии отлива. Южнее Горла более или менее постоянные поселения этого вида могут быть встречены лишь на слоевище сублиторальных водорослей в Кандалакшском заливе, в Бассейне и в северной части Онежского залива. На закрытых участках Поморского берега, вдоль Лямицкого, Летнего* и Зимнего берегов L. vincta если и встречается, то только единичными особями; в течение наших работ в 1948-1951 гг. она в этих местах не найдена вовсе.
* (На небольшом участке Летнего берега в районе острова Жижгина, где развиты пышные заросли ламинарий и других макрофитрв, имеются и постоянные поселения L. vincta.)
Таким образом, в отличие от прибрежных вод Восточного Мурмана, где этот моллюск в большем или меньшем количестве может быть обнаружен практически везде, распространение его южнее Горла ограничено лишь центральной частью водоема, а на огромном протяжении берегов Онежского и Двинского заливов постоянных и массовых поселений он не имеет.

Рис. 55. Кладки моллюсков. 1 - Кладки Littorina obtusaia на слоевище фукоида; 2 - Пелагические яйцевые капсулы L. littorea: а - сверху, б - вид сбоку; 3 - Кладки Lacuna palliduta: a - на слоевище Fucus serratus, б - увеличено; 4 - Кладки L. vincta: а - на слоевище ламинарии, б - увеличено; 5 - Кладки Margariles heiicina: a - на слоевище ламинарии, б - увеличено; 6 - Кладки Buccinum gronlandicum на камне и на раковине мидии; 7 - Кладка В. undatum на слоевище фукоида
При размножении L. vincta делает кладку (рис. 55, 4) на водорослях, из которой выходят планктонные личинки - велигер (Кузнецов 1948).
На Восточном Мурмане в том или ином местообитании единичные кладки могут быть обнаружены в любое время года, Тем не менее, в каждом местообитании обычно наблюдается четко выраженный сезонный цикл размножения. Например, на закрытых участках губ первые кладки обычно появляются в самый разгар зимы, в марте; особенно много их встречается с мая по август (табл. 53). Небольшое количество кладок появляется здесь еще в сентябре и даже в октябре. Выход из кладок планктонных личинок наблюдается с конца мая до начала сентября; кладки, появившиеся позднее этого срока, не развиваются и, очевидно, погибают. Молодые моллюски появляются среди прибрежных водорослей главным образом в конце августа - начале сентября.
| Месяцы | Всего | В том числе (в процентах) | |
| до стадии велигер |
на стадии велигер |
||
| I | 0 | 0 | 0 |
| II | 0 | 0 | 0 |
| III | 8 | 100 | 0 |
| IV | 62 | 100 | 0 |
| V | 141 | 95.8 | 4.2 |
| VI | 140 | 84.6 | 15.4 |
| VII | 121 | 84.4 | 15.6 |
| VIII | 102 | 82.0 | 18.0 |
| IX | 21 | 95.0 | 5.0 |
| X | 6 | 100 | 0 |
| XI | 0 | 0 | 0 |
| XII | 0 | 0 | 0 |
Таким образом, эмбрионы в кладках, отложенных в марте, заканчивают свое развитие приблизительно в течение 60 суток, а в отложенных летом - в течение 20-30 (по наблюдениям в садках). Превращение личинки в молодого моллюска завершается в более длительный срок (около 100 суток).
В западной части Кандалакшского залива Белого моря кладки L. vincta на ламинариях и зостере можно видеть с января по август, причем в значительно большем количестве, чем на Мурмане. Так например, на Мурмане на 10 растений ламинарий приходится в среднем не более 141 кладки, а в Кандалакшском заливе количество кладок достигает 344. Впрочем заметим, что в некоторые годы количество кладок на Восточном Мурмане в июне может достигать 740-1005 на каждые 10 слоевищ ламинарий (Кузнецов, 1948).
На литорали и в сублиторали восточной части Карельского берега кладки встречаются на стеблях зостеры и на слоевище других макрофитов почти исключительно зимой, с января по апрель или май, летний срок размножения здесь по существу отсутствует. С июня по август лишь очень редко можно обнаружить единичные кладки на слоевище сублиторальных макрофитов.
В отличие от прибрежных вод Западной Европы на Восточном Мурмане и в Белом море исчезновение кладок наблюдается в более теплое время года (табл. 54). Температурный диапазон, в котором происходит размножение беломорских особей, по существу включает в себя температурные диапазоны размножения как мурманских, так и западноевропейских особей; мурманские особи размножаются при температуре от 0 до 10, беломорские - от -1.5 до +16, а западноевропейские - от 1 до 18°. Таким образом, беломорские особи отличаются от западноевропейских способностью размножаться при отрицательных температурах, а от мурманских массовым размножением при значительно более высокой температуре.
| Место наблюдений | Месяцы | Средние месячные температуры воды, при которых происходит икрометание и развитие эмбрионов |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Южный берег Англии, район Плимута | + | + | + | + | + | * | * | * | * | * | * | * | 8-12 | Lebour, 1937 |
| О. Гельголанд | + | + | + | + | + | + | * | * | * | * | * | * | 1-14 | Ankel, 1936 |
| Восточный берег Дании, Зунд | * | * | + | + | + | + | + | * | * | * | * | * | 1-18 | Thorson, 1946 |
| Восточный Мурман, район губы Дальне-Зеленецкой |
* | * | + | + | + | + | + | + | + | * | * | * | 0-10 | Наши наблюдения |
| Белое море, район губы Гридиной | + | + | + | + | + | + | + | + | * | * | * | * | -1.5-+3.8 (+16) | Наши наблюдения |
| Западная часть Кандалакшского залива | + | + | + | + | + | + | + | + | * | * | * | * | -0.9-+16 | Наши наблюдения |
Примечание. + время икрометания и нахождения кладок.
На Восточном Мурмане каждая самка за период размножения делает от 6 до 25 кладок. По наблюдениям, в естественных условиях в 1939-1940 гг. каждая самка в среднем за период размножения делала от 9.0 до 11.6 кладок. Каждая кладка содержит здесь в среднем от 883 до 3447 эмбрионов*; среднее из всех наблюдений составляет 1802 (табл. 55). Кладки беломорских самок несколько уступают мурманским в отношении максимальных средних: наибольшее среднее количество зародышей в кладках мурманских особей достигает 3447, тогда как в Бассейне и заливах Белого моря - не более 2511. Однако общая средняя для беломорских кладок более высокая и достигает 2075.
* (В нашей статье о жизненном цикле этого вида (Кузнецов, 1948) сообщалось, что в 1939-1940 гг. кладки содержали от 320 до 1800 зародышей, в среднем 600. Сведения эти были основаны на сравнительно небольшом материале (просчитано количество зародышей в 25 кладках, полученных в садках) и поэтому могут дать ошибочное представление о действительности, если они не оказались следствием годовых колебаний разовой индивидуальной плодовитости.)
| Место наблюдений | Абсолютная амплитуда колебаний | Колебания средних в различных местообитаниях |
|
| минимум | максимум | ||
| Восточный Мурман | 280 | 6057 | 883-3447 |
| Белое море | |||
| Воронка и Горло | 729 | 4423 | 1006-3686 |
| Северный берег Бассейна и Кандалакшский залив |
785 | 3960 | 1985 |
| Северная часть Онежского залива | 931 | 4093 | 1772-2511 |
| Закрытые участки Карельского берега | 1294 | 2704 | 1775 |
| Среднее из всех наблюдений на Восточном Мурмане (216 кладок) | 1802 | ||
| Среднее из всех наблюдений в Белом море (81 кладка) | 2075 | ||
На Восточном Мурмане молодь, появившаяся осенью, в течение всей зимы довольно интенсивно растет (Кузнецов, 1948) и к весне достигает половой зрелости, имея при этом диаметр раковины в среднем 4.9 мм и вес 70 мг. Летом, как уже сказано, самки неоднократно делают кладки и с наступлением зимы в основной своей массе погибают. Лишь единичные особи сохраняются живыми и в течение зимы снова делают кладки. Продолжительность жизни моллюсков в естественных условиях не превышает 14-15 месяцев; только в исключительных случаях единичные особи живут до 17-18 месяцев. Заметим, что даже в условиях садка, где совершенно устраняется действие таких элиминирующих факторов, как нападение хищников, штормы и осеннее разрушение зарослей макрофитов, продолжительность жизни в 1939-1940 гг. не превышала 17-18, а в 1948-1949 гг. 20 месяцев; в этом возрасте моллюски имели диаметр раковины до 7.5 мм. В среднем диаметр раковины таких особей составлял 7.0 мм, а вес их был 105 мг. В естественных условиях средний размер раковины самых старших особей не превышал 5.8 мм (Кузнецов, 1948).
В Кандалакшском заливе Белого моря зимой мы сталкиваемся с такими условиями существования L. vincta, которые напоминают то, что имелось в наших мурманских садках. Под толщей ледяного припая здесь так же, как и в садках, полностью устранено элиминирующее влияние штормов и обусловленное ими катастрофическое разрушение зарослей макрофитов. Кроме этого, хотя зимнее охлаждение воды здесь и более продолжительное, чем на Мурмане, но в какой-либо мере резких колебаний температуры уже не бывает. Все это способствует лучшей выживаемости особей и, как следствие этого, вызывает увеличение продолжительности жизни, достигающей в таких условиях 2-2.5 лет. Наличием в составе зимней популяции значительного количества взрослых особей и обусловлено массовое появление кладок уже в январе, несмотря на то, что половая зрелость появившейся осенью молоди наступает лишь в возрасте 8-9 месяцев, т. е. не ранее апреля-мая будущего года. Для размножения этого вида условия Кандалакшского залива, по-видимому, менее благоприятны, и начиная с июля новых кладок здесь почти не появляется. У Карельского берега, как уже сказано, летний срок размножения практически отсутствует.
В общих чертах биологические особенности беломорских популяций представляются в следующем виде:
| * | Восточный Мурман |
Восточная часть Карельского берега |
Западная часть Кандалакшского залива |
| Продолжительность жизни (месяцы) | 14-15 | 7-8 | 24-30 |
| Возраст наступления половой зрелости (месяцы) | 7-9 | 3-5 | 8-9 |
| Диаметр раковины к концу жизни (мм) | 7.5 | 4-5 | 7.0 |
| Количество зародышей в кладке (среднее) | 1802 | 1775 | 1985 |
Таким образом, в Бассейне и заливах Белого моря имеется по крайней мере две популяции L. vincta, заметно отличающиеся по биологическим свойствам составляющих их особей как друг от друга, так и от популяции Восточного Мурмана.
Margarites helicina (Phipps) по характеру своего географического распространения должна быть отнесена, подобно предшествующим видам, к представителям бореальной фауны. Основным ее местообитанием на Восточном Мурмане, да и в Белом море служит пластинка каминарий, хотя нередко она заселяет заросли десмарестии и красных водорослей в сублиторали и некоторых фукоидов на литорали (Кузнецов, 1948); в Белом море к этому прибавляются еще и стебли зостеры. Заметим однако, что в Белом море наиболее массовые поселения этого моллюска обнаружены в прибрежных водах Бассейна и западной части Кандалакшского залива; у Карельского берега поселения малочисленны, а в южной половине Онежского залива и в Двинском заливе М. helicina, по-видимому, отсутствует вовсе.
При размножении М. helicina делает на водорослях кладку (рис. 55, 5), из которой выходит вполне сформировавшаяся молодь в ползающей стадии.
На Восточном Мурмане первые кладки, как правило, появляются в мае, иногда в апреле и даже в марте. Весь период икрометания продолжается здесь обычно с мая по август (табл. 56), а в некоторые годы до сентября. В кладках, отложенных в мае, эмбрионы достигают стадии вполне сформировавшейся молоди в июне, т. е. продолжительность их развития не превышает 30-40 суток; из позднее отложенных кладок молодь выходит уже через 20-25 суток. Вообще же период появления молоди продолжается на Мурмане с июня до сентября.
В Кандалакшском заливе Белого моря и у Карельского берега икрометание начинается только в июне и заканчивается в июле, т. е. продолжается всего лишь 1.5-2 вместо 4-7 месяцев на Мурмане. Продолжительность развития эмбрионов сокращается здесь до 15-20 суток.
| Месяцы | Всего | В том числе (в процентах) | |||
| яйца и зародыши до стадии велигер |
велигер | смешанные кладки |
вполне сформировавшая молодь |
||
| I-IV | Кладок не встречалось | ||||
| V | 111 | 78.1 | 21.9 | 0 | 0 |
| VI | 195 | 63.3 | 17.2 | 6.3 | 13.2 |
| VII | 230 | 30.0 | 35.0 | 14.9 | 20.1 |
| VIII | 189 | 8.1 | 19.8 | 12.0 | 60.1 |
| IX | 16 | 0 | 0 | 0 | 100 |
| X-XII | Кладок не встречалось | ||||
Количество зародышей в кладках беломорских особей, очевидно, несколько больше, чем мурманских (табл. 57). В различных местообитаниях Восточного Мурмана среднее количество зародышей в кладке составляло 103-137*, в Воронке Белого моря оно достигало 385, а в Бассейне и заливах колебалось от 137 до 172.
* (Описание кладки, данное в моей статье (Кузнецов, 1948), содержит ошибку. Дело в том, что описанная там кладка принадлежит не одной, а группе самок. Кладка одной самки М. helicina состоит всего лишь из одного комочка, но обычно несколько самок делают кладки в одном месте, в результате чего и образуются скопления, содержащие нередко 20 и даже более комочков.)
| Место наблюдений | Абсолютная амплитуда колебаний | Колебания средних в различных местообитаниях |
|
| минимум | максимум | ||
| Восточный Мурман | 42 | 358 | 103-137 |
| Белое море | |||
| Воронка | 85 | 761 | 385 |
| Горло | 62 | 119 | 81-98 |
| Северная часть Онежского залива | 63 | 275 | 137 |
| Полуоткрытые участки Карельского и Поморского берегов |
41 | 228 | 172 |
В заключение приводятся сведения о размерах раковины самых крупных особей в различных частях ареала*:
| * | Диаметр раковины самой крупной особи (в мм) |
* | Диаметр раковины самой крупной особи (в мм) |
| Гренландия | 8.5 | Воронка и Горло Белого моря | 9.0 |
| Исландия | 5.8 | Западная часть Кандалакшского залива | 11.0 |
| Шпицберген | 11.1 | Северная часть Онежского залива | 6.3 |
| Берингово море | 8.0 | Поморский берег и восточная часть Карельского берега |
5.0 |
| Охотское море | 14.1 | * | * |
| Южная Норвегия | 4.0 | * | * |
| Восточный Мурман | 8.0 | * | * |
* (Кроме наших наблюдений, сведения заимствованы еще из монографии Однера (Odhner, 1915).)
Как видно, особи из Онежского залива Белого моря превосходят по размерам лишь особей с побережий Исландии и южной Норвегии; особенно крупные моллюски встречаются в Охотском море, у Шпицбергена, в Кандалакшском заливе и в Воронке Белого моря. По-видимому, это обусловлено не ускорением роста, а увеличением продолжительности жизни. Мелкие размеры моллюсков в Онежском заливе и у Карельского берега являются следствием сокращения продолжительности жизни. На Восточном Мурмане половая зрелость моллюсков наступает в возрасте 11-12 месяцев, а максимальная продолжительность жизни составляет 22-23 месяца. В большинстве местообитаний Бассейна и заливов Белого моря половая зрелость наступает в возрасте 9-10 месяцев и особи не живут более одного года. Только в западной части Кандалакшского залива нередко можно встретить особей в возрасте до 4+ и имеющих размер до 11 мм. Значительное увеличение длительности жизни в данном случае часто способствует заметному увеличению численности, в результате чего основное местообитание этого вида - пластинка ламинарий бывает занята более плотными поселениями, чем на Мурмане.
Некоторые общие биологические особенности беломорских М. helicina представляются нам в следующем виде:
| * | Восточный Мурман |
Восточная часть Карельского берега |
Западная часть Кандалакшского залива |
| Продолжительность жизни (месяцы) | 22-23 | 11-12 | 48 и более |
| Возраст наступления половой зрелости (месяцы) | 11-12 | 9-10 | 9-10 |
| Диаметр раковины наибольшей особи (в мм) | 8.0 | 5.0 | 11.0 |
| Количество зародышей в одной кладке (среднее) | 103-137 | 172 | ? |
Таким образом, в Бассейне и заливах Белого моря имеется по меньшей мере две группы популяций этого вида, отличающихся друг от друга главным образом по размерам тела и продолжительности жизни.
Buccinum grönlandicum Chemnitz по характеру географического распространения должен быть отнесен к числу арктических, циркумполярных, видов. Он не опускается южнее побережья Норвегии, но довольно обычен в сибирских морях.
На Восточном Мурмане массовые поселения В. gronlandicum. имеются почти всюду на камнях и между камнями в нижней половине литоральной зоны. Зимние скопления моллюсков бывают особенно плотными, и на 1 м2 дна в это время года собирается до 1180 особей. В результате того, что летом особи занимают значительно больший, чем зимой, экологический ареал, плотность поселений в это время года не превышает 150-440 экз. с биомассой 230-870 г/м2 дна. В Воронке Белого моря поселения этого вида столь же многочисленны, как и на Восточном Мурмане.
Южнее Горла распространение В. gronlandicum ограничено, главным образом, берегами Бассейна и прилегающих к нему участков Онежского и Кандалакшского заливов. Массовых поселений, подобных мурманским, здесь не обнаружено.
При размножении В. gronlandicum делает кладку (рис. 55, 6), из которой выходит вполне сформировавшаяся молодь в ползающей стадии.
На Восточном Мурмане самки делают кладки в ноябре (табл. 58) и развитие эмбрионов заканчивается лишь на будущее лето: выход молоди наблюдается в июле-сентябре. В восточной части Карельского берега Белого моря значительное количество кладок встречается на камнях и литоральных фукоидах в течение лета.. Кладки, собранные в июле, содержали в себе эмбрионов на самых начальных стадиях развития, тогда как на Мурмане в это время года наблюдался уже выход молоди, а имеющиеся еще в кладках эмбрионы заканчивали свое развитие. Очевидно, что в Белом море происходит серьезное изменение времени размножения этого вида с перемещением периода икрометания на лето.
| Месяцы | Количество кладок (в %) с различными стадиями развития эмбрионов | |||
| яйца | эмбрионы до стадии велигер |
велигер | вполне сформировавшаяся молодь |
|
| I | 100 | 0 | 0 | 0 |
| II | 50 | 17 | 33 | 0 |
| III | 0 | 50 | 50 | 0 |
| IV | - | - | - | - |
| V | 0 | 0 | 100 | 0 |
| VI | 0 | 0 | 100 | 0 |
| VII | 0 | 0 | 38 | 62 |
| VIII | 0 | 0 | 0 | 100 |
| IX | 0 | 0 | 50 | 50 |
| X | кладок не найдено | |||
| XI | 100 | 0 | 0 | 0 |
| XII | 0 | 100 | 0 | 0 |
Продолжительность жизни на Восточном Мурмане и в Белом море одинакова и в обоих случаях не превышает 7+ лет. Самые крупные особи на Восточном Мурмане имеют высоту раковины 39.5, а в Белом море 37.6 мм. Средняя высота раковины в возрасте 6+ лет в первом случае составляет 34.0, а во втором 33.2 мм. Таким образом, в какой-либо мере заметных отличий в скорости роста и продолжительности жизни особей этого вида в Белом море по сравнению с Восточным Мурманом пока не обнаруживается.
Висcinum undatum L. образует массовые поселения у берегов Западной Европы, где наносит существенный вред рыболовству, повреждая попавшую в сети рыбу. Восточный Мурман и Белое море служат лишь северо-восточной границей распространения этого бореального вида, и численность его здесь сравнительно небольшая. На Восточном Мурмане В. undatum в теплое время года служит обычным представителем литоральной фауны с плотностью поселений не более 10 особей с биомассой до 98.9 г/м2. С наступлением осенних холодов моллюски уходят в сублитораль, где и живут до следующей весны. На литорали Бассейна и заливов Белого моря этот вид встречается редко и в большинстве случаев живет только в сублиторали, на глубине до 15-20 м и более.
При размножении самки этого вида делают кладку (рис. 55, 7), из которой выходит вполне сформировавшаяся молодь в ползающей стадии. На Восточном Мурмане свежеотложенные кладки встречаются с апреля по август, а выход молоди замечен в сентябре-октябре. Каждая кладка составлена из 57-135 отдельных капсул, содержащих от 1 до 22 зародышей каждая, в среднем 5.8. Кладки, добытые в южной половине Онежского залива Белого моря, состояли из 51-66 капсул и в каждой капсуле было от 1 до 12 эмбрионов, в среднем 5.7. Кладка далеко не всегда делается одной самкой, нередко несколько самок делают общую кладку, поэтому приведенный материал не может дать правильного представления о размерах индивидуальной плодовитости беломорских и мурманских особей. Количество же эмбрионов в одной капсуле может свидетельствовать лишь о том, что репродуктивные способности тех и других весьма сходные.
На Восточном Мурмане особи живут до 12+ лет и к концу жизни имеют раковину высотой до 72.2-81.2 мм; в Бассейне и заливах Белого моря продолжительность жизни и максимальные размеры тела уменьшаются. Даже в наиболее благоприятном в этом отношении районе - северной части Онежского залива - особи не живут более 10-11 лет, а раковины их не бывают более 66.7 мм высотой. В Кандалакшском заливе продолжительность жизни не превышала 6 + , а высота раковины самой крупной особи 41.7 мм, а у Летнего берега (Двинский залив) максимальный возраст был 7 + , а высота раковины 55.8 мм.
Муtilus edulis L. является одним из наиболее широкораспространенных прибрежных моллюсков. В Северной Атлантике он доходит на юге до Средиземного моря в Европе и до северной Каролины в Америке, а в прибрежных водах Шпицбергена, Земли Франца Иосифа, в Карском и других сибирских морях, по-видимому, отсутствует. Литоральные поселения мидий имеют одну из своих границ на Восточном Мурмане, далее к востоку, в Чошской губе и у Новой Земли, они уже уходят в сублитораль до глубины 30 м (Гурьянова, 1930). В Белом море мидии в одинаковой мере широко распространены и образуют массовые поселения, как на литорали, так и в сублиторали.
На литорали Восточного Мурмана максимальная биомасса мидий на отвесных скалах полуоткрытого участка литорали достигает 28.8 кг/м2 при количестве особей более 70 тыс. (Матвеева, 1948а; Кузнецова, Матвеева, 1948). На илисто-песчанистых пляжах закрытых участков литорали биомасса не превышает здесь 14.3 кг/м2 при 4600 экз. На Белом море плотности поселений значительно меньшие, по нашим данным, не более 5-6 кг/м2.
Уменьшение плотности поселений и биомассы мидий на беломорской литорали вполне компенсируется развитием обширных сублиторальных повелений, практически отсутствующих на Восточном Мурмане (Солдатов, 1934; Паленичко, 1943; 1947). Сублиторальные мидиевые банки особенно сильно развиты в Двинском заливе и вдоль Летнего берега на глубинах до 10-15 м, где биомасса достигает 12 кг/м2. Вдоль Поморского и Карельского беретов и у берегов многочисленных островов Онежского залива сублиторальные поселения мидий занимают обширные площади, но биомасса их не превышает здесь 0.4-0.6 кг/м2. В Кандалакшском заливе и в Бассейне Белого моря массовых сублиторальных поселений мидий не обнаружено, но отдельные особи и небольшие группы их встречаются там на глубине до 265 м (Книпович, 1906а; Дерюгин, 1928). На такие большие глубины мидии заносятся с литорали льдами или на оторвавшихся водорослях.
Таким образом, существенным отличием беломорских мидий от мидий прибрежных вод Восточного Мурмана является резкое расширение экологического ареала и образование массовых поселений не только на литорали, но и в сублиторали. Получается интересный ряд:
Восточный Мурман - массовые поселения, как правило, только на литорали.
Юго-восточная часть Баренцева моря - массовые поселения только в сублиторали.
Белое море - массовые поселения на литорали и в сублиторали, отдельные особи живут на самых больших глубинах.
Известно, что у берегов Западной Европы мидии также обитают не только на литорали, но и в сублиторали, где поселяются совместно с устрицами и нередко даже вытесняют их (Möbius, 1877).
Размножение беломорских мидий изучено пока еще недостаточно, можно лишь отметить, что вымет половых продуктов заканчивается здесь значительно раньше, в более короткий срок и протекает при более высоких температурах, чем на Мурмане (табл. 59). В июле, а нередко уже в конце июня гонады оказываются пустыми, тогда как на Восточном Мурмане вымет половых продуктов растягивается на период с мая до сентября (Матвеева, 1948). Заметим, что на Мурмане вымет половых продуктов происходит при наиболее низких температурах по сравнению со всеми другими частями ареала этого вида, а на Белом море это происходит при таких же температурах, как и у берегов Англии, Франции, Дании и других европейских стран.
| Место наблюдений | Месяцы | Средние месячные температуры воды, при которых происходит икрометание и развитие эмбрионов |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Восточный берег Северной Америки, район Новой Шотландии |
* | * | * | + | + | + | + | + | + | * | * | * | 8-12 | Field, 1909, 1923 |
| Ирландское море, берег Ланкашира | * | * | * | * | + | + | + | * | * | * | * | * | 10-15 | Scott, 1901 |
| Южный берег Англии, район Плимута | + | + | + | + | + | * | * | * | * | * | * | * | 8-13 | Matthews, 1913 |
| Северный берег Франции | * | + | + | + | + | + | + | + | + | * | * | * | 7-17 | Field, 1909 |
| О. Гельголанд и западный берег Дании | * | * | + | + | + | + | + | + | + | + | * | * | 3-19 | Havinga, 1929 |
| Восточный берег Дании, Зунд | * | * | * | * | + | + | * | * | * | * | * | * | 7-14 | Thorson, 1946 |
| Фиорды Дании, Айс-фиорд, Лим-фиорд | * | * | * | * | + | + | * | * | * | * | * | * | 10-15 | Runnström, 1928 |
| Южный берег Норвегии, район Бергена | * | * | + | + | + | + | * | * | * | * | * | * | 4-11 | Thorson, 1946; Runnström, 1928 |
| Восточный Мурман, район губы Дальне-Зеленецкой |
* | * | * | + | + | + | + | * | * | * | * | * | 1-6 | Матвеева, 1948 |
| Белое море, Бассейн и заливы | * | * | * | * | * | + | + | * | * | * | * | * | 6-16 | Наши наблюдения |
Примечание. + время икрометания.
Наибольшая продолжительность жизни мидий на Восточном Мурмане составляет 13+ лет, а в Белом море обычно 17+ лет*. В соответствии с этим находится и размер раковины: среди литоральных мидий Восточного Мурмана самые крупные имеют высоту раковины 52.7, а в Белом море 54.2 мм; сублиторальные мидии Восточного Мурмана могут иметь высоту раковины 62.5, а беломорские 75.2 мм (табл. 60).
* (А. И. Савилов (1954), специально занимавшийся изучением биологии мидий Кандалакшского залива, считает, что здесь они имеют продолжительность жизни до 25 лет. В наших сборах столь великовозрастных особей не встречалось; лишь в одном случае была найдена мидия в возрасте 22 лет. Если прав Савилов, то длительность жизни беломорских мидий по сравнению с мидиями североморскими увеличивается в 3-4 раза, а максимальные размеры тела уменьшаются в 2-2.5 раза.)
| Характер местообитания | Возраст (годы) | Максимальный возраст (годы) |
||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | самая крупная особь |
||
| Открытые берега мелких островов | ||||||||||
| Восточный Мурман Длина раковины (в мм) |
5.5 | 9.3 | 13.7 | 18.0 | 22.0 | 25.9 | 29.4 | 32.2 | 52.7 | 9+ |
| Интенсивность роста (в %) | * | 69.1 | 47.3 | 31.4 | 22.2 | 27.7 | 13.5 | 9.5 | * | |
| Северная часть Онежского залива Длина раковины (в мм) |
5.1 | 9.3 | 13.5 | 16.5 | 18.9 | 20.9 | 22.0 | 23.8 | 28.6 | 9+ |
| Интенсивность роста (в %) | * | 82.3 | 45.2 | 22.2 | 14.5 | 10.6 | 5.3 | 8.2 | * | |
| Средняя часть Онежского залива Длина раковины (в мм) |
5.6 | 10.0 | 14.0 | 18.0 | 21.6 | 26.8 | 29.1 | 30.7 | 34.2 | 9+ |
| Интенсивность роста (в %) | * | 78.6 | 40.0 | 28.6 | 20.0 | 24.1 | 8.6 | 5.5 | * | |
| Южная часть Онежского залива Длина раковины (в мм) |
6.3 | 10.9 | 15.1 | 19.2 | 22.9 | 27.4 | 30.8 | 32.0 | 37.5 | 8+ |
| Интенсивность роста (в %) | * | 73.0 | 38.5 | 27.2 | 19.3 | 19.6 | 12.4 | 3.9 | * | |
| Открытые участки материкового берега | ||||||||||
| Восточный Мурман Длина раковины (в мм) |
5.1 | 8.4 | 12.8 | 17.1 | 21.6 | 24.8 | 28.6 | 32.5 | 42.0 | 12+ |
| Интенсивность роста (в %) | * | 64.7 | 52.3 | 33.5 | 26.3 | 14.8 | 15.3 | 13.6 | * | |
| Воронка Белого моря Длина раковины (в мм) |
5.7 | 9.8 | 14.3 | 19.0 | 21.8 | 24.8 | 27.2 | 28.4 | 36.0 | 13+ |
| Интенсивность роста (в %) | * | 71.9 | 45.9 | 32.9 | 15.8 | 13.8 | 9.7 | 4.4 | * | |
| Северный берег Бассейна Длина раковины (в мм) |
5.5 | 9.8 | 13.7 | 17.2 | 20.4 | 22.2 | 24.7 | 27.0 | 28.4 | 8+ |
| Интенсивность роста (в %) | * | 78.5 | 39.8 | 25.5 | 18.6 | 8.8 | 11.3 | 9.3 | * | |
| Северный берег Кандалакшского залива Длина раковины (в мм) |
6.2 | 13.0 | 18.5 | 22.6 | 25.4 | 27.0 | - | - | 31.0 | 6+ |
| Интенсивность роста (в %) | * | 109.7 | 42.3 | 22.2 | 12.3 | 6.3 | -4 | - | * | |
| Каменистые россыпи на закрытых участках | ||||||||||
| Восточный Мурман (губа Дальне-Зеленецкая) Длина раковины (в мм) |
4.7 | 8.8 | 13.7 | 18.7 | 23.2 | 26.9 | 30.2 | 33.0 | 51.6 | 12+ |
| Интенсивность роста (в %) | * | 87.2 | 55.7 | 36.5 | 24.1 | 15.9 | 12.3 | 9.3 | * | |
| Кандалакшский залив (губы Падан и Шарапиха, подножие Крестовой горы) Длина раковины (в мм) |
5.9 | 11.5 | 16.7 | 22.0 | 26.2 | 31.4 | 35.3 | 37.2 | 52.0 | 10+ |
| Интенсивность роста (в %) | * | 94.9 | 45.2 | 31.7 | 19.1 | 19.8 | 11.8 | 5.4 | * | |
| Восточная часть Карельского берега (губа Гридина) Длина раковины (в мм) |
6.0 | 12.0 | 18.2 | 23.8 | 28.4 | 32.6 | 35.6 | 39.2 | 54.2 | 17+ |
| Интенсивность роста (в %) | * | 100.0 | 51.7 | 30.8 | 19.2 | 14.8 | 9.2 | 10.1 | * | |
| Северная половина Онежского залива (о-ва Русский Кузов и Жижгин) Длина раковины (в мм) |
6.5 | 13.1 | 19.8 | 25.6 | 30.4 | 38.7 | 35.9 | 37.4 | 45.8 | 10+ |
| Интенсивность роста (в %) | * | 101.5 | 51.1 | 29.3 | 19.0 | 10.9 | 6.5 | 4.2 | * | |
| Южная половина Онежского залива (губа Колежма, Сумская губа) Длина раковины (в мм) |
5.4 | 11.0 | 13.5 | 16.9 | 20.2 | 22.7 | 25.2 | 26.4 | 32.0 | 9+ |
| Интенсивность роста (в %) | * | 103.7 | 22.7 | 25.2 | 19.5 | 12.4 | 11.0 | 4.8 | * | |
| Илисто-песчанистые пляжи в кутовых участках губ | ||||||||||
| Восточный Мурман (губа Дроздовка) Длина раковины (в мм) |
5.4 | 9.4 | 13.2 | 16.7 | 20.0 | 23.2 | 26.2 | 28.8 | 44.2 | 13+ |
| Интенсивность роста (в %) | * | 74.1 | 40.4 | 26.5 | 19.8 | 17.0 | 12.0 | 9.9 | * | |
| Восточная часть Карельского берега (губа Гридина) Длина раковины (в мм) |
4.1 | 10.3 | 16.8 | 21.5 | 24.3 | 27.8 | - | -8 | 32.3 | 7+ |
| Интенсивность роста (в %) | * | 151.2 | 63.2 | 28.02 | 13.0 | 14.0 | - | - | * | |
| Различные грунты ниже линии отлива | ||||||||||
| Восточный Мурман Длина раковины (в мм) |
5.6 | 10.8 | 17.4 | 25.7 | 32.7 | 39.9 | 44.7 | 48.7 | 62.5 | 11+ |
| Интенсивность роста (в %) | * | 92.8 | 61.1 | 47.7 | 27.2 | 22.0 | 12.0 | 8.9 | * | |
| Южная часть Горла Белого моря (между д. Пялица и Пулонга) Длина раковины (в мм) |
8.3 | 15.3 | 22.2 | 29.1 | 35.0 | 39.0 | 42.0 | 45.3 | 51.5 | 13+ |
| Интенсивность роста (в %) | * | 84.3 | 43.8 | 21.1 | 20.3 | 11.4 | 7.7 | 7.9 | * | |
| Северная часть Онежского залива (около о. Жижгин) Длина раковины (в мм) |
9.1 | 18.9 | 29.8 | 33.9 | 39.5 | 45.8 | 48.8 | 51.2 | 75.2 | 12+ |
| Интенсивность роста (в %) | * | 107.7 | 57.1 | 13.8 | 16.5 | 15.3 | 6.6 | 4.9 | * | |
| Средняя часть Онежского залива (около о-вов Б. Жужмуй и Кондостров) Длина раковины (в мм) |
8.0 | 15.4 | 21.9 | 27.6 | 33.0 | 36.6 | 42.0 | 44.1 | 64.0 | 12+ |
| Интенсивность роста (в %) | * | 92.5 | 42.2 | 26.0 | 19.6 | 10.9 | 14.8 | 5.0 | * | |
| Южная часть Онежского залива (около о-вов Тонкая Осинка, Турья Луда и Березовец) Длина раковины (в мм) |
7.2 | 13.4 | 19.5 | 24.9 | 29.4 | 32.7 | 35.2 | 37.3 | 50.0 | 8+ |
| Интенсивность роста (в %) | * | 86.1 | 45.6 | 27.7 | 18.1 | 11.2 | 7.6 | 6.0 | * | |
| Средняя часть Горла у Зимнего берега (около д. Инцы) Длина раковины (в мм) |
5.4 | 9.1 | 12.5 | 15.2 | 17.0 | 19.8 | 21.8 | 23.8 | 29.0 | 9+ |
| Интенсивность роста (в %) | * | 58.5 | 37.4 | 21.3 | 11.8 | 16.5 | 10.1 | 9.2 | * | |
Характерной чертой беломорских мидий в целом служит образование многих групп популяций, отличающихся друг от друга такими важнейшими биологическими свойствами, как продолжительность жизни, скорость роста и размеры тела. Следует особенно подчеркнуть, что на закрытых участках губ и заливов, где условия существования более суровые, продолжительность жизни не уменьшается, как это можно было бы ожидать, а чаще всего увеличивается.
Таким образом, кратко характеризуя биологические особенности беломорских мидий, необходимо отметить следующее:
- Экологический ареал значительно расширяется, включая в себя не только различные местообитания литорали, но и сублитораль. В этом отношении беломорские мидии сходны с мидиями прибрежных вод Западной Европы и вообще умеренных широт. Плотность поселений в Белом море заметно снижается, хотя общая численность увеличивается благодаря значительному расширению занимаемых площадей.
- Период икрометания завершается в более сжатые сроки по сравнению с подавляющим большинством других частей ареала и протекает при значительно более высоких, чем на Мурмане, температурах, весьма близких к умеренным широтам.
- На первом году жизни беломорские мидии в большинстве местообитаний крупнее, чем на Мурмане, но к концу жизни примерно одинаковы с ними. Для Белого моря характерно образование множества более или менее самостоятельных популяций, отличающихся друг от друга многими биологическими свойствами, такими, как скорость роста, длительность жизни и др. Максимальная длительность жизни в Белом море более высокая, чем на Мурмане и по сравнению с любыми другими частями ареала.
- Многие беломорские популяции этого вида, занимающие открытые участки побережий, составлены особями, угнетенными в отношении роста и размеров тела, а особи закрытых участков побережий и сублиторали наоборот отличаются ускорением роста и более крупными размерами.
Сardium ciliatum Fabr. служит представителем арктической фауны и широко распространен во всех северных морях, образуя нередко весьма плотные поселения. Например, в Печорском районе Баренцева моря биомасса достигает 365 и в фиордах Восточной Гренландии 134 г/м2 (Зенкевич, 1926, 1947; Spärck, 1933, Броцкая, Зенкевич, 1939). В Северной Атлантике южная граница ареала проходит в районе Исландии и мыса Код.
В Баренцевом море этот моллюск распространен по всей акватории и может быть встречен на любых глубинах, хотя и обнаруживает предпочтение к глубинам 30-80 м. В Белом море он обычно глубже 30-35 м не опускается.
У берегов Восточного Мурмана в местообитаниях С. ciliatum отрицательных температур практически никогда не бывает, а в период максимального прогрева температура воды может подниматься до 6-8°. Амплитуда сезонных колебаний температур здесь (на глубине около 50 м) составляет обычно не более 5-7°. К юго-западу от Шпицбергена на глубине 365 м температура воды и все сопутствующие ей факторы подвержены весьма незначительным сезонным колебаниям; отрицательных температур никогда не бывает, а годовая амплитуда температурных колебаний едва ли превышает здесь 3-4°. В районе Земли Франца Иосифа и в Маточкином Шаре особи этого вида большую часть года живут при отрицательных температурах воды, сменяющихся положительными (около 1.5°) только на короткий период лета. Амплитуда сезонных колебаний температуры в местообитаниях моллюска здесь очень незначительна и обычно не превышает 2.5-3°. В районе Шпицбергена амплитуда сезонных колебаний температуры также незначительна, но отличие в условиях существования состоит в том, что там относительное постоянство термического режима сопутствовало довольно высокой средней температуре, тогда как здесь оно сопутствует крайне низкой средней температуре. В Печорском районе Баренцева моря, а также в районе ВорОнкн Белого моря сильное зимнее охлаждение (до -1.5°) сменяется довольно сильным летним прогреванием (до 5-7°). Амплитуда сезонных колебаний температуры в этих районах заметно увеличивается и становится такой же, как в прибрежных водах Восточного Мурмана. Но разница в температурных условиях существования С. ciliatum обнаруживается очень существенная: в прибрежных водах Восточного Мурмана этот моллюск почти никогда не встречается с отрицательными температурами, а в Печорском районе и в Воронке Белого моря ежегодно подвергается их воздействию в течение нескольких месяцев. В Онежском заливе Белого моря, в местах массовых поселений С. ciliatum температура воды колеблется от -1.5 зимой до +12° летом; амплитуда сезонных колебаний температуры в этом случае является наибольшей из всех перечисленных местообитаний.
К сожалению, нам почти ничего неизвестно об особенностях размножения этого моллюска в каждом местообитании, и поэтому мы можем рассмотреть лишь некоторые особенности роста и продолжительности жизни, что сравнительно легко определяется по годовым кольцам на раковине (табл. 61).
| Место наблюдений | Количество изученных особей |
Длина раковины (в мм) | Максимальный возраст (годы) |
||||||||
| 1 год | 2 года | 3 года | 4 года | 5 лет | 6 лет | 7 лет | 8 лет | самая крупная особь |
|||
| Прибрежные воды Восточного Мурмана. Глубины до 150 м | 145 | 7.2 | 12.1 | 17.0 | 22.2 | 29.5 | 30.5 | 33.3 | 36.4 | 45.5 | 10 |
| Район Воронки Белого моря. Глубины до 100 м | 11 | 4.4 | 10.1 | 15.3 | 21.1 | 27.3 | 32.3 | 36.6 | 40.0 | 61.4 | 30 |
| Район острова Колгуева. Глубины до 40 м | 44 | 6.7 | 11.2 | 14.6 | 19.2 | 23.8 | 27.7 | 30.1 | 31.2 | 53.0 | 31 |
| Район Маточкина Шара. Глубины до 75 м | 18 | 4.2 | 6.8 | 9.0 | 11.2 | 13.6 | 15.6 | 16.7 | 19.2 | 37.5 | 26 |
| Район Шпицбергена. Глубины до 365 м | 6 | 7.4 | 10.8 | 14.7 | 17.5 | 20.5 | 21.6 | 25.8 | 26.9 | 44.4 | 33 |
| Кандалакшский залив. Глубины до 30 м | 34 | 4.2 | 7.9 | 11.4 | 13.6 | 16.8 | 21.2 | 24.6 | 27.0 | 36.0 | 12 |
| Онежский залив. Глубины до 30 м | 423 | 3.5 | 6.5 | 9.0 | 11.5 | 14.3 | 16.4 | 18.4 | 20.5 | 33.6 | 13 |
Вполне достоверные сведения о скорости роста раковины этого вида получены нами лишь для прибрежных вод Восточного Мурмана и для Онежского залива Белого моря. Правильное представление о скорости роста и продолжительности жизни получено также для Печорского района Баренцева моря и для Кандалакшского залива Белого моря. Для всех остальных пунктов приводятся лишь предварительные сведения, характеризующие порядок величин.
Наиболее высокой скоростью роста отличаются особи из прибрежных вод Восточного Мурмана и из района Воронки Белого моря, но продолжительность жизни в прибрежных водах Восточного Мурмана не превышает 10 лет, тогда как в Воронке Белого моря она достигает 30 лет. Столь же высокая продолжительность жизни свойственна и моллюскам из Печорского района, но скорость роста здесь значительно снижена, поэтому моллюски не достигают таких крупных размеров, как в районе Воронки. Сравнивая между собой три перечисленных участка Баренцева моря, нетрудно понять, что в этом случае более сильное летнее прогревание в прибрежной зоне способствует ускорению роста, но более резкие сезонные колебания температуры и других факторов отрицательно сказываются на продолжительности жизни. В районе Воронки летнее прогревание становится слабее, чем в прибрежных водах Восточного Мурмана, но и амплитуды колебаний условий внешней среды несколько снижаются, это благоприятно сказывается и на скорости роста и на выживаемости; в результате моллюски достигают здесь в 3 раза большего возраста и значительно более крупных размеров. Некоторое возрастание амплитуры колебаний факторов внешней среды в Печорском районе вызывает снижение скорости роста, оставляя продолжительность жизни пока неизменной. Дальнейшее увеличение амплитуды колебаний условий внешней среды, что имеет место в Онежском заливе Белого моря, в дополнение к снижению скорости роста вызывает и сокращение продолжительности жизни; это приводит к измельчанию особей. Если в Баренцевом море длина раковины наиболее крупных особей составляла 61.4 мм, то в Белом море особи крупнее 36 мм в настоящее время не встречаются. Таким образом, увеличение амплитуды сезонных колебаний факторов внешней среды приводит в конечном итоге к измельчанию особей, что происходит главным образом в результате уменьшения продолжительности жизни.
Существование моллюсков при постоянных низких температурах, несмотря на сравнительно небольшую амплитуду их сезонных колебаний, неблагоприятно сказывается прежде всего на скорости роста. Скорость роста снижается настолько сильно, что, несмотря на довольно высокую продолжительность жизни, моллюски остаются мелкими. Примером этого могут служить особи из района Маточкина Шара. Таким образом, в таких случаях измельчание особей происходит главным образом в результате снижения скорости роста. Иными словами, на северо-востоке Баренцева моря и в Белом море обитают почти одинаково мелкие особи С. ciliatum, по в первом случае это измельчание является результатом снижения скорости роста, а во втором - уменьшения продолжительности жизни.
Приведенный здесь материал позволяет в качестве биологических особенностей беломорских кардиум отметить следующее: общее сокращение ареала, резкое сокращение длительности жизни и снижение скорости роста, начиная с первого же года жизни.
Serripes ( = Cardium) grönlandicum (Chemn.), кроме Баренцева и Белого морей, широко распространен на мелководьях всех арктических морей. В Северной Атлантике он встречен у берегов Норвегии, Исландии, Ян-Майена, Гренландии и вдоль побережья Америки до мыса Код.
В Баренцевом море этот вид образует более или менее массовые поселения только в восточной части на илистых и илисто-песчанистых грунтах Печорского района и района Воронки Белого моря. Здесь, на глубинах 20-80 м, обитают наиболее крупные особи и биомасса их достигает 135 г/м2 при 50 экз. У берегов и в губах Восточного Мурмана S. grönlandicum встречается довольно часто, но массовых поселений уже не образует. Любопытно, что К. М. Дерюгин в своей монографии о фауне Кольского залива (1915) указывал на существование больших поселений моллюсков на глубине 18-50 м. Возможно, что в связи с длительным периодом потепления эти поселения нацело исчезли. В Белом море небольшие скопления S. grönlandicum обычны в губах Кандалакшского залива на глубине 30-40 м; мелкие единичные особи могут быть встречены и в Онежском заливе.
Размножение этого вида не изучалось, и поэтому мы можем рассмотреть лишь вопросы, связанные с ростом и продолжительностью жизни.
Наиболее высокая скорость роста и наиболее крупные размеры свойственны особям из юго-восточной части Баренцева моря; продолжительность жизни здесь также достигает максимума (табл. 62). Как известно, эта часть моря характеризуется сравнительно резкими сезонными колебаниями температуры: зимние отрицательные температуры воды сменяются здесь довольно хорошим летним прогреванием. В отличие от этого, для прибрежных вод Шпицбергена характерно слабое летнее прогревание и сильное зимнее охлаждение; амплитуда сезонных колебаний здесь небольшая, но и общее количество тепла, которым могут воспользоваться обитатели этих вод, очень ограничено. В результате, подобно тому как это было показано у Cardium ciliatum, скорость роста моллюсков уменьшается, но продолжительность жизни все еще остается высокой, что приводит к заметному уменьшению размеров самых крупных особей; например, если в районе мыса Святой Нос длина раковины самой крупной особи достигала 94.5 мм, то в прибрежных водах Шпицбергена особи крупнее 60.2 мм не обнаружены. Интересно, что для этого арктического вида, более "арктические" условия прибрежных вод Шпицбергена оказались менее благоприятны, чем более умеренные условия юго-восточной части Баренцева моря. Одновременно с этим, для Serripes grönlandicum еще менее благоприятными оказались и близкие к "бореальным" условия жизни в прибрежных водах Восточного Мурмана. Правда, в губах Восточного Мурмана в течение первых 3-4 лет жизни особи растут даже с несколько повышенной скоростью по сравнению с особями из юго-восточной части Баренцева моря, но уже с 4-5 года жизни скорость их роста резко падает, что вместе с сокращением продолжительности жизни приводит к сильному уменьшению размеров самых крупных особей.
| Место наблюдений | Количество изученных особей |
Длина раковины (в мм) | Максимальный возраст (годы) |
||||||||
| 1 год | 2 года | 3 года | 4 года | 5 лет | 6 лет | 7 лет | 8 лет | самая крупная особь |
|||
| Прибрежные воды Восточного Мурмана. Глубина 25 м | 13 | 8.6 | 13.2 | 17.4 | 21.3 | 23.1 | 25.3 | 27.5 | 29.8 | 46.7 | 10 |
| К северу от м. Святой Нос. Глубины 88-90 м | 37 | 5.4 | 10.5 | 16.3 | 22.4 | 29.6 | 37.5 | 44.4 | 50.6 | 94.5 | 38 |
| Прибрежные воды Шпицбергена. Глубины 24-50 м | 21 | 6.3 | 9.8 | 13.9 | 18.5 | 25.2 | 28.4 | 30.9 | 34.0 | 60.2 | 33 |
| Губы Кандалакшского залива Белого моря. Глубины 6-20 м | 9 | 6.5 | 11.7 | 19.1 | 23.0 | 26.2 | - | - | - | 28.0 | 5 |
| Южная и западная (прибрежная) часть Онежского залива. Глубины 7-17 м | 22 | 2.9 | 5.1 | 6.5 | 8.5 | * | * | * | * | 9.3 | 4 |
В Кандалакшском заливе Белого моря скорость роста остается высокой, но продолжительность жизни ныне живущих особей в нашем материале не превышала 5 лет, в результате чего этот вид представлен здесь сильно измельчавшими особями. Крайнюю степень угнетения S. grönlandicum можно наблюдать в Онежском заливе, т. е. в условиях наиболее высоких амплитуд сезонных колебаний температуры и других факторов. По сборам в 1937 г., длина раковины самых крупных особей не превышала здесь 9.3 мм, а продолжительность жизни 4 лет; иными словами, по сравнению с оптимумом в юго-восточной части Баренцева моря в Онежском заливе продолжительность жизни и максимальные размеры тела уменьшаются в десять раз.
В последние годы в связи с климатическими колебаниями возрастной состав популяции этого вида здесь несколько изменился. По сборам в течение 1946-1952 гг. (Кудерский, 1958), наиболее крупные особи в большинстве местообитаний Онежского залива имели длину раковины уже до 20 мм, а в одном случае даже 35 мм. Максимальный возраст моллюсков достигал 8 лет. Статья Л. А. Кудерского написана в остро полемическом тоне и, может быть, поэтому в ней допущены неточности и искажения. Прежде всего необходимо отметить, что указание на находку в Онежском заливе особи, имеющей раковину длиной 73.8 мм и возраст 15+ лет просто не соответствует действительности. Факт этот проверен мною лично. В коллекциях Беломорской Биологической Станции такой экземпляр действительно имеется, но без какого-либо указания о месте его находки. Из беседы с сотрудниками Станции выяснилось, что более вероятно предположить, что он добыт не в Онежском заливе, а в Воронке Белого моря. Утверждение Л. А. Кудерского, что этот вид является обычным в Онежском заливе и имеет там крупные размеры, опровергается его же сообщением о величине максимальной биомассы в. 5.1 г/м2. Такая величина биомассы свидетельствует лишь о господстве здесь молодых особей и редкой встречаемости взрослых. Совершенно очевидно, что достаточно, попасть в дночерпатель только одной особи, указываемых Л. А. Кудерским размеров, как биомасса их на квадратный метр на много превзойдет величину 5.1 г. Впрочем, статья Л. А. Кудерского, наряду с некоторыми интересными фактами, содержит и множество других совершенно недопустимых погрешностей, главной из которых является то, что при анализе материала им далеко не всегда отделялись пустые раковины и створки от живых особей.
Сущность же биологических особенностей беломорских представителей этого вида, даже если принять за основу данные Л. А. Кудерского, остается прежней: резкое сокращение длительности жизни, снижение скорости роста и уменьшение численности.
Суprina islandica L. служит обычным представителем бореальной фауны и довольно широко распространена на мелководьях и в прибрежных водах Баренцева и Белого морей. Кроме этого, она хорошо известна в фиордах и вообще в прибрежных водах Норвегии, Фарерских островов и Исландии, в прибрежных водах западной части Балтийского моря, у берегов Дании и Франции. Вдоль восточного берега Северной Америки она встречается от залива Лаврентия до мыса Гаттераса. В прибрежных водах Шпицбергена, Земли Франца Иосифа и далее на се.вер С. islandica очевидно отсутствует. Таким образом, прибрежные воды Восточного Мурмана и Белое море служат краевой частью ареала этого бореального вида.
В пределах своего ареала С. islandica населяет мягкие грунты на глубинах от 10 до 120 м. В некоторых случаях, например в губах Западного Мурмана, значительные поселения могут встречаться даже на глубине 1-2 м (Лейбсон, 1939). Большую часть своей жизни моллюски проводят, зарывшись глубоко в грунт, что и делает их мало доступными для обычных орудий лова (драги, тралы и даже дночерпатели). Лишь в конце зимы, в феврале-апреле, животные поднимаются на поверхность грунта, в результате чего взятые в это время года драги и тралы приносят большое число особей. Так обстоит дело в прибрежных водах Восточного Мурмана. В отличие от этого в Белом море С. islandica, по-видимому, живет на поверхности грунта в течение круглого года и поэтому в большом количестве добывалась нами в летние месяцы. Для сравнения можно указать, что в прибрежных водах Восточного Мурмана наши тралы в период с мая по декабрь никогда не приносили одновременно более 24 молодых и 6 взрослых особей, тогда как в Онежском заливе около Жужмуйских островов траление на глубине 30 м принесло более 780 особей различных возрастов: за одно траление здесь было добыто больше материала, чем за все время работ в прибрежных водах Восточного Мурмана.
В юго-восточной части Баренцева моря наибольшая биомасса достигает 26 г/м2 при 145 экз. (Броцкая, Зенкевич, 1926). Соотношение между весом и количеством особей показывает, что господствующее положение здесь занимает молодь. Далее к западу, в Мотовском заливе (Лейбсон, 1939), наибольшая биомасса уже составляет 427 г/м2 при 46 экз. Если в первом случае средняя длина раковины составляла 8, то здесь она равна уже 32 мм. Еще далее к западу, в фиордах Фарерских островов биомасса увеличивается до 815 г/м2 при 15 экз.; это соответствует средней длине раковины моллюсков около 60 мм. В фиордах Исландии биомасса достигает своего максимума - 1221.5 г/м2 при 40 экз.; средняя длина раковины моллюска здесь может быть около 48 мм (Spärck, 1929, 1937).
В Кандалакшском заливе Белого моря единичные особи встречаются в губах на глубине около 30-40 м. Численность С. islandica здесь настолько ограничена, что в работе по количественному учету фауны в прибрежных водах Кандалакшского залива (Гурвич, Иванов, 1939) лишь упоминается о встречаемости этого вида, но никаких количественных сведений не дается. В северной половине Онежского залива, на глубине около 30 м, наоборот, поселения моллюска, как уже сказано, очень многочисленны.
Об особенностях размножения беломорских С. islandica пока ничего неизвестно, поэтому мы ограничимся рассмотрением лишь некоторых особенностей роста и продолжительности жизни.
Самые крупные особи, отличающиеся наиболее высокой скоростью роста, в районе наших исследований населяют открытые части прибрежных вод Восточного Мурмана (табл. 63). Эти особи занимают такие местообитания, где отрицательных температур никогда не бывает, а сезонные колебания температуры и многих других факторов приближаются к минимуму. Амплитуда температурных колебаний составляет здесь около 4-5° и ниже 1.4° вода не охлаждается. В проливах между островами и материком при наличии сильных приливо-отливных течений из-за близости суши амплитуда температурных колебаний может возрастать до 5.5°. Сходство условий существования в открытых частях прибрежных вод Восточного Мурмана и в проливах между островами и материком служит причиной и близкого сходства скорости роста и продолжительности жизни обитающих здесь С. islandica. Как видно, продолжительность жизни в этих двух случаях составляет 26-28 лет, а средняя длина раковины в возрасте 8 лет 47.8-49.1 мм.
| Место наблюдений | Количество изученных особей |
Длина раковины (в мм) | Максимальный возраст (годы) |
||||||||
| 1 год | 2 года | 3 года | 4 года | 5 лет | 6 лет | 7 лет | 8 лет | самая крупная особь |
|||
| Открытые части Баренцева моря в районе к северу от м. Териберского. Глубины 60-100 м | 12 | 5.8 | 9.4 | 15.0 | 22.2 | 26.5 | 31.8 | 40.0 | 49.1 | 92.0 | 28 |
| Прибрежные воды Восточного Мурмана на участках с сильными приливо-отливными течениями. Глубины 40-50 м | 35 | 5.5 | 9.3 | 15.1 | 21.1 | 26.5 | 32.0 | 41.1 | 47.8 | 81.9 | 26 |
| Губы Восточного Мурмана. Глубины 25-60 м | 41 | 3.3 | 5.7 | 8.1 | 11.9 | 14.5 | 17.1 | 19.2 | 19.8 | 30.7 | 11 |
| Северная половина Онежского залива. Глубина 30 м | 730 | 3.6 | 5.7 | 7.8 | 10.1 | 12.3 | 14.6 | 16.9 | 19.5 | 34.5 | 17 |
| Губы Кандалакшского залива. Глубины 13-25 м | 56 | 3.5 | 5.9 | 8.8 | 11.6 | 15.0 | 18.2 | 20.5 | 23.8 | 49.2 | 28 |
В губах Восточного Мурмана в некоторые годы в местообитаниях моллюска уже могут быть отрицательные температуры до -1°, в летнее время прогревание увеличивается настолько, что амплитуда сезонных колебаний температуры достигает уже 6-8°. В результате скорость роста, продолжительность жизни и размеры тела животных уменьшаются. Продолжительность жизни составляет не более И лет, а длина раковины в возрасте 8 лет в среднем 19.8 мм и вообще особи крупнее 30.7 мм здесь не встречались.
В средней части Онежского залива, как это уже сказано, условия жизни подвержены наиболее резким сезонным колебаниям, что, по-видимому, и служит причиной некоторого сходства между моллюсками, обитающими здесь и в губах Восточного Мурмана: продолжительность жизни в Онежском заливе не превышает 17 лет, а длина - раковины в возрасте 8 лет составляет в среднем 19.5 мм. В Кандалакшском заливе в местообитаниях С. islandica сезонные колебания температуры и сопутствующих ей факторов в значительной мере сокращаются главным образом за счет уменьшения летнего прогревания. В результате этого продолжительность жизни становится такой же, как в наиболее, благоприятных условиях Баренцева моря (28 лет), но скорость роста продолжает оставаться сильно замедленной, и средняя длина раковины в возрасте 8 лет составляет лишь 23.8 мм, т. е. в два раза меньше, чем в Баренцевом море.
Таким образом, в качестве основных отличий беломорских С. islandica от баренцевоморских мы можем пока назвать сокращение ареала и численности, а также уменьшение скорости роста, начиная с первого же года жизни.
Ракообразные
Anonyx nugax (Phipps) наиболее массовые поселения образует в юго-восточной части Баренцева моря и в прилегающих частях Полярного Бассейна, Карского и Гренландского морей; не менее массовые поселения имеются и в Белом море.
По характеру питания A. nugax может быть причислен к типичным падальщикам, пожирающим не только трупы животных, но и попавшую в сети еще живую рыбу, успевая в течение одной ночи полностью ее уничтожить или сильно повредить. Массовое количество аноникса в Белом море и вред, причиняемый им рыболовству, известны давно. Вот например, что писали о нем в конце XVIII в. (Звягин, 1794:161): "Сии копшаки столь ядовиты, что не только одних сельдей изъедают, но равномерно точат увязших в ловчие сети тюленей и семги, и вящему удивлению сосут и переедают сельдяные сети, которые уже впредь для лова становятся негодными". Для сбора материала по этому виду мы и воспользовались его способностью в массовом количестве нападать на мясо или рыбу. Сбор материала производился при помощи специальных ловушек (Кузнецов, Матвеева, 1948), выставляемых в различных пунктах на тот или иной промежуток времени. Об относительной численности этого вида можно судить по количеству особей, добываемых в течение суток. По наблюдениям в сентябре-октябре, наибольший суточный улов в Кандалакшском заливе достигал 119 112 особей, а на Мурмане в это время года суточные уловы не превышали 3526. Средние суточные уловы были в это время года в прибрежных водах Восточного Мурмана 1486, в Воронке Белого моря 2156, в Горле Белого моря 2874, в Двинском заливе 1299, в Кандалакшском заливе 19 406 и в Онежском заливе 6003 особей.
Высокая численность A. nugax в Белом море сопровождается и значительно более крупными размерами тела (табл. 64). В прибрежных водах Восточного Мурмана осенью длина тела самых крупных особей не превышает 18, а в Белом море достигает 40 мм. За все время наших наблюдений у берегов Восточного Мурмана длина тела самой крупной особи едва достигала 24 мм, а в Белом море нередко в массовом количестве встречаются особи с длиной тела 44-46 мм.
| Длина тела (в мм) |
Губа Дальне-Зеленецкая, октябрь, глубина 6 м |
У дер. Кашкаренцы, октябрь, глубина 6 м |
Губа Падан, средняя часть, сентябрь |
О. Воль-остров, октябрь, глубина 16 м |
|
| глубина 6 м | глубина 20 м | ||||
| 36-38 | 0 | 0 | 0 | 0 | 5 |
| 34-36 | 0 | 0 | 0 | 2 | 10 |
| 32-34 | 0 | 0 | 0 | 4 | 13 |
| 30-32 | 0 | 0 | 0 | 2 | 21 |
| 28-30 | 0 | 0 | 0 | 0 | 5 |
| 26-28 | 0 | 0 | 0 | 0 | 15 |
| 24-26 | 0 | 0 | 0 | 13 | 33 |
| 22-24 | 0 | 0 | 0 | 9 | 51 |
| 20-22 | 0 | 0 | 0 | 14 | 49 |
| 18-20 | 0 | 8 | 0 | 38 | 44 |
| 16-18 | 10 | 16 | 39 | 69 | 29 |
| 14-16 | 20 | 47 | 435 | 334 | 159 |
| 12-14 | 10 | 98 | 286 | 253 | 152 |
| 10-12 | 80 | 146 | 133 | 194 | 173 |
| 8-10 | 730 | 250 | 5 | 42 | 139 |
| 6-8 | 150 | 255 | 29 | 18 | 29 |
| 4-6 | 0 | 180 | 13 | 8 | 71 |
| 2-4 | 0 | 0 | 0 | 0 | 2 |
В прибрежных водах Восточного Мурмана продолжительность жизни особей составляет около 3, а в Белом море она увеличивается до 4 и даже 6 лет. Половая зрелость в обоих морях наступает в возрасте около полутора лет; в прибрежных водах Мурмана самки вскоре после первого размножения погибают, а в Белом море каждая самка в течение своей жизни может размножаться до 4 раз.
В юго-западной части Баренцевого моря откладывание самками яиц в пределах всей популяции заканчивается за период с января по апрель. Отложив в выводковую сумку яйца, самки отходят от берегов и больше сюда уже никогда не возвращаются. Молодь рождается в открытом море и в середине лета подходит к берегам. В Белом море икрометание может происходить и фактически происходит в течение круглого года, хотя массовое рождение молоди наблюдалось только в августе.
В прибрежных водах Восточного Мурмана размножающиеся самки имеют длину тела от 12.5 до 23.5 мм и вынашивают от 15 до 201 зародыша; в среднем каждая самка в разные годы вынашивает от 51 до 105 зародышей.
В бассейне и заливах Белого моря длина тела размножающихся самок в общем колебалась от 11 до 46 мм, а индивидуальная плодовитость от 15 до 950 зародышей. Впервые размножающиеся самки, т. е. в одинаковом возрасте с мурманскими, имели длину тела от 11 до 22.5 мм и вынашивали от 15 до 78 зародышей, в среднем 43. Возможно, что впервые размножающиеся самки действительно здесь менее плодовиты, чем в прибрежных водах Восточного Мурмана, но, к сожалению, беломорский материал на этот счет довольно ограничен.
Сравнение индивидуальной плодовитости самок при одинаковых размерах тела показывает, что репродуктивные способности мурманских особей более высокие, чем беломорских:
| Длина тела (в мм) |
Восточный Мурман |
Белое море |
| 40-46 | - | 406 |
| 30-40 | - | 303 |
| 24-30 | - | 286 |
| 20-24 | 103 | 60 |
| 16-20 | 83 | 54 |
| до 16 | 58 | 26 |
Таким образом, беломорские А. nugax отличаются от мурманских более крупными размерами тела, большей продолжительностью жизни, более высокой плодовитостью и резко возросшей численностью. Весьма возможно, что в Белом море образуется две группы популяций этого вида. Одна такая группа населяет верхние слои воды и отличается от мурманских сравнительно мелкими размерами тела, пониженной плодовитостью и высокой численностью. По аналогии с некоторыми из ранее изученных видов можно ожидать, что особям этой группы свойственны и более раннее наступление половой зрелости и общее сокращение продолжительности жизни. Популяции этой группы занимают всю акваторию Онежского залива, а также мелководья в других районах Белого моря, где нередко они уже смешиваются с особями другой группы популяций. Эта группа популяций населяет преимущественно большие глубины Бассейна и Кандалакшского залива, где господствуют малоизменчивые низкие температуры; осенью особи этих популяций нередко поднимаются на мелководья и здесь смешиваются с особями первой группы популяций. Особи популяций, населяющих глубины Белого моря, отличаются от мурманских очень высокой численностью, крупными размерами тела, возросшей продолжительностью жизни и более высокой плодовитостью.
Rhachotropis aculeata (Lepechin) является представителем арктической фауны и широко распространен в морях северного полушария, предпочитая участки, более или менее удаленные от материковых берегов. В Баренцевом море этот вид образует по крайней мере три биологически разнокачественных популяции (рис. 56):
I. Западная баренцевоморская популяция занимает район, ограниченный на юге 30 и 39 меридианами, далее на север полоса поселений постепенно суживается и доходит до 75° с. ш. Глубины здесь колеблются от 136 до 365 м, а сезонные и региональные колебания температуры лежат в пределах от 0.10 до 3.80°. Самки этой популяции достигают поло вой зрелости и размножаются в возрасте 10-11 месяцев и имеют продолжительность жизни до 2 лет. Весь цикл размножения (икрометание, вынашивание зародышей и рождение молоди) начинается и заканчивается в течение зимы. Самые крупные особи имеют длину тела до 38 мм.
Размножающиеся самки имеют длину тела от 22.5 до 32.9 мм (в среднем 26.3 мм) и плодовитость от 16 до 111, в среднем каждая самка вынашивает 62 зародыша.
Рис. 56. Распределение различных биологических групп Rhachotropis aculeata в Баренцевом и Белом морях и в прилегающих частях Полярного Бассейна. I - западная баренцевоморская, II - восточная баренцевоморская, III - северная и IV - беломорская популяции
II. Восточная баренцевоморская популяция заселяет главным образом юго-восточный угол Баренцева моря, расселяясь здесь на глубинах 75-92 м, при сезонных и региональных температурных колебаниях от -2.0 до -0.29°. В отличие от западной популяции размножение восточной популяции протекает в общем в течение круглого года, но при этом наблюдается два максимума: зимний и летний. Самки зимнего поколения (т. е. родившиеся зимой) достигают половой зрелости й размножаются в возрасте около полутора лет и дают начало летнему поколению. Самки летнего поколения (т. е. родившиеся летом) достигают половой зрелости в том же возрасте и дают начало зимнему поколению. Таким образом, самостоятельность зимнего ж летнего поколений не сохраняется, и они постоянно переходят друг в друга. Общая продолжительность жизни, по-видимому, составляет здесь около 2.5 лет, и самые крупные особи достигают в длину 39 м. Размножающиеся самки имеют длину тела от 27.1 до 39.0 мм (в среднем 32.3 мм) и плодовитость от 52 до 241, в среднем каждая самка вынашивает 117 зародышей.
III. Северная популяция расселяется на глубинах от 60 м и глубже в районах, непосредственно прилегающих к Шпицбергену и Земле Франца Иосифа. Особи этой популяции встречаются не только в северной части Баренцева моря, но и в Гренландском, Норвежском и Карском морях, а также в Полярном Бассейне. В местообитаниях этой популяции характерны почти неизменные температуры от -1.8 до -1.0°. Размножение протекает круглый год, как и у особей восточной популяции. Важным биологическим отличием особей северной популяции является увеличение продолжительности жизни (до 4 лет и больше?) и крупные размеры тела: самые крупные из них достигают длины 44 мм.
Длина тела размножающихся самок колеблется от 32.7 до 44.0 мм (в среднем 39.8 мм), а индивидуальная плодовитость от 25 до 162, составляя в среднем 95 зародышей. Как видно, несмотря на резкое увеличение размеров тела самок этой популяции, их индивидуальная плодовитость значительно сократилась.
IV. Белое море населено самостоятельной популяцией, отличающейся целым рядом биологических свойств от перечисленных трех популяций Баренцева моря. Беломорская популяция населяет глубины преимущественно 55-56 м и прижимается к северному берегу Бассейна. Сезонные и региональные колебания температуры здесь достигают довольно высокой амплитуды: температура от -1.5 зимой может подниматься до 3.8-4.0° летом. Ни одна из трех баренцевоморских популяций не живет при столь широких колебаниях температуры. По характеру жизненного цикла особи беломорской популяции сходны с особями западной баренцевоморской популяции, но отличаются от них крайне быстрым ростом и высокой плодовитостью. При одинаковой продолжительности жизни (около 2 лет) с особями западной баренцевоморской популяции особи беломорской популяции достигают длины тела 44 мм, т. е. такой же, как особи северной популяции за 4 года жизни. Размножающиеся самки в Белом море имеют длину тела от 32.5 мм до 43.7 мм (в среднем 37.2 мм) и вынашивают от 66 до 401 зародыша, в среднем 222.
Сравнение индивидуальной плодовитости самок перечисленных четырех популяций (табл. 65) наглядно свидетельствует о высокой биологической активности представителей этого вида в Белом море.
| Длина тела (в мм) |
Баренцевоморская | Северная | Беломорская | |
| западная | восточная | |||
| 42-44 | - | - | 138 | 326 |
| 40-42 | - | - | 114 | 312 |
| 38-40 | - | 188 | 92 | 240 |
| 36-38 | - | 181 | 25 | 253 |
| 34-36 | - | 158 | - | 211 |
| 32-34 | 102 | 122 | 65 | 101 |
| 30-32 | 98 | 97 | - | - |
| 28-30 | 63 | 86 | - | - |
| 26-28 | 68 | 84 | - | - |
| 24-26 | 56 | - | - | - |
| 22-24 | 42 | - | - | - |
Примечание. Индивидуальная плодовитость здесь и во всех других случаях соответствует среднему количеству зародышей, вынашиваемых одной самкой.
По частоте встречаемости и по количеству особей, добываемых за одно траление, судя по имевшимся в нашем распоряжении материалам, на первое место следует поставить восточную баренцевоморскую популяцию, за ней идет беломорская, а затем уже северная и западная баренцевоморская.
Таким образом, основными биологическими отличиями беломорских особей служат высокая скорость роста, крупные размеры тела и возросшая плодовитость.
Gammarus locusta (L.) в отличие от предшествующих видов является типичным литоральным обитателем, широко распространенным вдоль побережий морей Северной Атлантики, в том числе Баренцева и Белого. Лишь в очень редких случаях возможны находки особей этого вида ниже линии отлива, на глубинах до 20 м.
Основным местообитанием гаммаруса служит узкое пространство между нижней поверхностью лежащих на литорали камней и подстилающим грунтом. На литорали Мурманского побережья животные обычно всю свою жизнь проводят в одном и том же местообитании, тогда как в Белом море наличие ледяного припая позволяет им в течение зимы занимать всю литоральную зону и только на лето уходить под камни.
На литорали Восточного Мурмана G. locusta живет при сезонных и региональных колебаниях температуры воды от -3 до +13°, а в Белом море от -2 до +22°.
Продолжительность жизни особей в обоих морях в общем одинакова и колеблется от 7-10 до 14 месяцев, при наступлении половой зрелости в возрасте от 1 до 7 месяцев.
На Мурмане первые икряные самки появляются в октябре-декабре; икрометание почти непрерывно продолжается до августа будущего года. Рождение молоди начинается в самый разгар зимы, в феврале, и заканчивается в августе. Весь процесс размножения протекает при температуре воды от -3 до +13°. В Белом море (южнее Горла) икряные самки впервые появляются уже в сентябре или в первые дни октября; особенно интенсивное размножение протекает зимой подо льдом. Заканчивается икрометание не позднее первой половины июля. Рождение молоди начинается в январе и заканчивается в июле. Весь процесс размножения протекает здесь при температуре воды от -2 до 22°.
Продолжительность периода размножения у G. locusta в Баренцевом и Белом морях столь же велика, как и у берегов Дании, но там оно протекает при более высоких температурах (табл. 66).
| Место наблюдений | Месяцы | Средние месячные температуры воды, при которых происходит икрометание и инкубация |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Gammarus locusta | ||||||||||||||
| Побережье Дании, Ниборг-фиорд | + | + | + | + | + | + | + | + | + | + | * | * | 1-18° | Blegvad, 1922 |
| Восточный Мурман, район губы Дальне-Зеленецкой |
+ | + | + | + | + | + | + | + | * | + | + | + | -0.2+10 | Наши наблюдения |
| Белое море, Бассейн и заливы | + | + | + | + | + | + | + | + | + | + | + | + | -1.5+16 | Наши наблюдения |
| G. marinus | ||||||||||||||
| Южный берег Англии, район Плимута | + | + | + | + | + | * | * | * | * | * | + | + | 7-12 | Cheng, 1942 |
| Восточный Мурман, район губы Дальне-Зеленецкой |
+ | + | + | + | + | + | * | * | * | * | * | * | -0.2+6 | Наши наблюдения |
| Белое море, Бассейн и заливы | * | * | + | + | + | + | + | + | + | * | * | * | -1.5+16 | Наши наблюдения |
Примечание. + время икрометания и инкубации.
Плодовитость и размеры тела размножающихся самок изменяются в весьма широких пределах. На литорали Восточного Мурмана длина тела размножающихся самок колеблется от 9 до 29 мм, а среднее количество зародышей (в разных местообитаниях и в разное время года) от 48 до 63. В Воронке и в северной части Горла Белого моря размножающиеся самки имеют длину тела от 14 до 29 мм и в среднем каждая самка вынашивает (в разных местообитаниях и в разное время года) от 53 до 113 зародышей. В Бассейне и заливах Белого моря обнаруживаются уже явные признаки угнетения этого вида: длина тела размножающихся самок колеблется от 11 до 23 мм, а средняя плодовитость (в разных местообитаниях и в разное время года) от 44 до 58.
Интересные различия между мурманскими и беломорскими особями обнаруживаются при сравнении индивидуальной и относительной плодовитости летом и зимой (табл. 67).
| Место наблюдения | n | Длина тела (мм) M ± m |
Индивидуальная плодовитость M ± m |
Корреляция между длиной тела и индивидуальной плодовитостью (r ± mr) |
Количество зародышей на 1 г живого веса |
| Июнь-август | |||||
| Открытые берега Воронки и Горла Белого моря | 341 | 21.1±0.2 | 113±2 | +0.48±0.04 | 475 |
| Открытые берега Восточного Мурмана | 126 | 19.1±0.3 | 54±2 | +0.63±0.05 | 286 |
| Закрытые берега Восточного Мурмана | 335 | 16.6±0.1 | 63±2 | +0.66±0.03 | 508 |
| Открытые берега Бассейна и заливов Белого моря | 210 | 16.1±0.2 | 58±2 | +0.62±0.04 | 513 |
| Закрытые берега Бассейна и заливов Белого моря | 108 | 15.0±0.2 | 41±2 | +0.54±0.04 | 471 |
| Январь-апрель | |||||
| Открытые берега Восточного Мурмана | 136 | 19.4±0.3 | 48±3 | +0.27±0.08 | 245 |
| Закрытые берега Восточного Мурмана | 473 | 16.6±0.1 | 48±1 | +0.41±0.04 | 387 |
| Открытые и полуоткрытые берега Бассейна и заливов Белого моря |
100 | 16.6±0.2 | 52±2 | +0.57±0.07 | 473 |
Средние размеры тела мурманских и беломорских размножающихся самок в разное время года практически одинаковы, тогда как плодовитость на Мурмане зимой уменьшается, а в Белом море, наоборот, увеличивается. Относительная плодовитость беломорских зимних самок может быть в 1.5-2 раза боле высокой, чем у мурманских.
Подобное положение является следствием того, что зима для беломорских особей оказывается более мягкой, чем для мурманских, благодаря теплоизолирующему действию ледяного припая.
Наиболее крупные и наиболее плодовитые самки, как это видно, живут на литорали Воронки и Горла Белого моря.
Таким образом, особи Бассейна и заливов Белого моря в целом отличаются от особей Восточного Мурмана, Воронки и северной части Горла Белого моря меньшей численностью, более мелкими размерами тела и меньшей плодовитостью. Особенно резка эти биологические особенности выявляются в течение лета, зимой же мурманское особи в ряде случаев уступают беломорским.
Крупные размеры тела, высокая плодовитость и высокая численность особей в Воронке и в северной части Горла Белого моря обусловлены, по-видимому, увеличением продолжительности жизни и более поздним наступлением половой зрелости по сравнению с тем, что наблюдается на Мурмайе и тем более в Бассейне и заливах Белого моря.
Gammarus marinus Leach. подобно G. locusia является типичным литоральным обитателем, встречающимся ниже линии отлива лишь в виде крайне редкого исключения. Местообитания обоих видов очень сходны, и иногда их популяции перемешиваются как на Мурмане, так и в Белом море.
Продолжительность жизни особей различных поколений и популяций G. marinus в обоих морях примерно одинакова и колеблется от 5-6 до 12-14 месяцев, при наступлении половой зрелости в возрасте от 2 до 8 месяцев.
На литорали Восточного Мурмана икрометание начинается в декабре-феврале и заканчивается в июне, т. е. протекает в сравнительно холодное время года. Молодь рождается в течение периода с марта по август, но массовое появление молоди замечено в марте-апреле и в июне-августе. В отличие от этого на литорали Бассейна, Кандалакшского и Онежского заливов откладывание яиц начинается очевидно только в марте и заканчивается в августе. Рождение молоди протекает здесь с апреля по сентябрь. На Восточном Мурмане размножение наблюдается почти в те же сроки, что и у берегов Англии (Cheng, 1942), но при значительно более низких температурах. На Белом море время размножения, как это видно, несколько меняется и занимает все лето; икрометание и развитие эмбрионов здесь может происходить при самых высоких температурах (табл. 66).
Длина тела размножающихся самок на литорали Восточного Мурмана в разное время года и в разных местообитаниях колеблется от 9.0 до 18.3 мм, а средняя индивидуальная плодовитость - от 9 до 37 зародышей. Сходные данные получены и для Воронки и северной части Горла Белого моря, где длина тела размножающихся самок была от 10.0 до 18.0 мм, и в различных местообитаниях плодовитость была от 15 до 29 зародышей.
Южнее Горла длина тела размножающихся самок и их плодовитость заметно уменьшаются: длина тела колеблется от 7.4 до 15.9 мм, а средняя плодовитость в различных местообитаниях изменяется от 12 до 16 зародышей.
Так же как и у летних G. locusta, индивидуальная и относительная плодовитость беломорских самок G. marinus в большинстве размерных групп заметно уступает тому, что можно наблюдать у мурманских самок. Лишь у самых мелких беломорских самок относительная плодовитость увеличивается, и они в этом отношении превосходят мурманских (табл. 68). В целом же беломорские самки уступают мурманским не только по плодовитости, но и по размерам тела.
| Длина тела (в мм) |
Восточный Мурман | Белое море | ||||
| индивидуальная | относительная | Воронка и Горло | Бассейн и заливы | |||
| индивидуальная | относительная | индивидуальная | относительная | |||
| 16-17 | - | - | 32 | 340 | - | - |
| 15-16 | - | - | 22 | 286 | - | - |
| 14-15 | 24 | 374 | 22 | 343 | - | - |
| 13-14 | 18 | 337 | 19 | 356 | 12 | 230 |
| 12-13 | 14 | 318 | 16 | 363 | 14 | 318 |
| 11-12 | 17 | 488 | 12 | 344 | 12 | 344 |
| 10-11 | 12 | 480 | - | - | 10/td> | 400 |
| 9-10 | 10 | 588 | - | - | 11 | 646 |
| 8-9 | - | - | - | - | 9 | 962 |
| 7-8 | - | - | - | - | 6 | 667 |
Pandalus annulicorni s Leach. в Баренцевом море образует массовые поселения вдоль открытых берегов Восточного Мурмана и обычно не идет далее 8-10 миль от берега, предпочитая глубины около 60 м. К востоку от Горла Белого моря не встречается (Дерюгин, 1928). Незначительные поселения его могут быть обнаружены в устьевых участках губ Восточного Мурмана на глубине 25-40 м. В закрытых губах и на меньших глубинах не был найден ни разу. У берегов Норвегии этот вид опускается на глубины до 650 м (Wollebaek, 1908).
В Белом море массовые поселения P. annulicornis обнаружены в северных частях Онежского и Двинского заливов на глубинах от 30 до 60 м, а в Горле и в Воронке на глубинах 30-70 м. Единичные особи 4 этого вида встречаются почти повсеместно. Плотность поселений в Баренцевом и Белом морях, по-видимому, одинакова: в обоих морях за одно траление добывалось не более 100-150 особей. Массовые скопления особей наблюдаются .осенью и зимой: в другое время года их не бывает.
В отличие от P. annulicomis другой представитель этого рода - P. borealis Kroyer живет преимущественно в открытых частях Баренцева моря, а в Белом море встречается очень редко (рис. 57), поэтому о нем мы здесь говорить не будем.
Рис. 57. Распространение Pandalus borealis (○) и P. annulicornis (●) в Баренцевом и Белом морях
Рассмотрим некоторые биологические свойства особей разных популяций P. annulicornis.
Известно, что у этого вида имеется смена полов (Jagersten, 1936; Leloup, 1936). Например, у западного побережья Швеции около 60% молоди в возрасте 1 1/2 лет функционирует в качестве самцов, а через год, т. е. в возрасте 2 1/2 лет, они превращаются в самок и откладывают икру на плеоподы (так называемые "вторичные" самки). Остальная часть молоди, минуя стадию самца, уже в возрасте 1 1/2 лет, непосредственно превращается в самок и принимает участие в размножении. Предельная продолжительность жизни самок составляет здесь 4 года, а длина их тела не превышает 72 мм. Откладывание яиц происходит в ноябре, а выход личинок в мае-июне.
В прибрежных водах Восточного Мурмана также имеются "первичные" и "вторичные" самки, причем, первых обычно бывает больше, чем вторых (в среднем около 35 "вторичных" и 65% "первичных"). "Вторичные" самки размножаются здесь только один раз в жизни, после чего погибают, имея общую продолжительность жизни около 3 лет. Длина тела самых крупных самок достигает 78 мм. Откладывание яиц наблюдалось в ноябре-декабре, а выход личинок в планктон в апреле. В Белом море икряные самки в значительном количестве встречаются с конца сентября или с начала октября. Нередко размножающиеся самки попадаются здесь даже в июле и августе.
В Бассейне и заливах Белого моря "вторичных" самок, по-видимому, не бывает, а "первичные" живут до 3-4 лет, размножаются до 2-3 раз в жизни. Самки, размножающиеся по второму и третьему разу, откладывают икру не осенью, как впервые размножающиеся, а в июле. Тем не менее, инкубация этих столь рано отложенных яиц заканчивается только к весне, т. е. одновременно: с отложенными осенью.
В различных участках прибрежных вод Восточного Мурмана средняя длина тела размножающихся самок колеблется от 52.3 мм на глубинах менее 100 м до 62.9 мм в районе мыса Святой Нос. Индивидуальная плодовитость соответственно изменяется в пределах от 1162 до 1397. В. Бассейне и заливах Белого моря средняя длина тела размножающихся самок составляет 56.3 мм, а плодовитость 1356 (табл. 69).
| Место наблюдений | n | Длина тела (в мм) | Плодовитость | Корелляция между длиной тела и плодовитостью (r±mr) |
||||
| M±m | σ±mσ | V±mv | M±m | σ±mσ | V±mv | |||
| Восточный Мурман. Прибрежные воды. Глубины до 100 м | 207 | 52.3±0.5 | 7.2±0.3 | 13.8±0.6 | 1162±32 | 472±23 | 40.6±2.2 | +0.71±0.03 |
| Открытая часть прибрежных вод. Глубины более 100 м | 169 | 58.2±0.6 | 7.8±0.5 | 13.4±0.8 | 1271±44 | 524±33 | 41.5±2.8 | +0.54±0.06 |
| То же, район м. Святой Нос. Глубины 102-107 м | 34 | 62.9±1.1 | 6.4±0.7 | 10.2±1.2 | 1397±102 | 610±71 | 43.4±6.0 | +0.71±0.06 |
| Бассейн и залив Белого моря. Глубины 10-60 м | 25 | 56.3±1.5 | 7.3±1.0 | 13.0±1.9 | 1356±140 | 698±99 | 51.5±9.0 | -0.05 |
Отсутствие смены полов в Белом море приводит к тому, что самцы там имеют длительность жизни, равную с самками, и достигают длины тела 70 мм, в то время как, например, у берегов Швеции они крупнее 60 мм не встречались.
Добавим еще, что в Баренцевом море самая мелкая икряная самка имела длину тела 34, а в Белом море 47 мм. Поскольку возраст наступления половой зрелости в обоих случаях одинаков, т. е. около 2 лет, то это свидетельствует об ускорении роста беломорских особей.
Таким образом, беломорские представители этого типично бореального вида отличаются от баренцевоморских: отсутствием смены полов и "вторичных" самок, заметным ускорением роста в течение по крайней мере первых 2 лет жизни и увеличением плодовитости по сравнению с "первичными" самками Баренцева моря. Характерно также увеличение продолжительности жизни и многократное размножение самок. В заключение заметим, что материал по беломорским представителям этого вида пока еще не может считаться достаточным и дальнейшие наблюдения и сборы могут еще многое уточнить, а частично и изменить. Spirontocaris spinus (Sowerby) является представителем арктической фауны и широко распространен в Баренцевом море. В зоне встречи теплых и холодных водных масс, т. е. между 32 и 38 меридианами он поднимается почти до 76° с. ш., массовые же поселения его тянутся узкой полосой вдоль всего побережья Восточного Мурмана. В районе мыса Святой Нос эта полоса массовых поселений раздваивается: с одной стороны, она идет в Горло и в южные районы Белого моря, а с другой, через Канинско-Колгуевское мелководье переходит к берегам Новой Земли, вдоль которых снова поднимается до 76° с. ш. (рис. 58). Небольшие поселения S. spinus имеются в прибрежных водах Земли Франца Иосифа и Шпицбергена.
В губах Восточного Мурмана S. spinus встречается не всегда и более обычен в открытом море на расстояние от 2 до 10 миль от берега на глубинах до 210 м.
В Белом море небольшие поселения этого вида обнаружены в Кандалакшском и Онежском заливах на глубинах от 12 до 40 м. Несколько увеличивается его численность в Бассейне, особенно в районе, непосредственно примыкающем к Горлу, на глубинах от 30 до 70 м.
Продолжительность жизни основной массы особей в Баренцевом море очень немногим превышает один, год: особи, закончившие размножение весной, живут до июня-июля и погибают, уступая место молодому поколению. Незначительное число особей доживает до осени и размножается вторично. Интересно, что у всех самок, закончивших размножение весной, в течение июня и июля идет интенсивное развитие гонад, но по неизвестной причине подавляющее большинство их вскоре погибает и в размножении не участвует.
Откладывание яиц на плеоподы в Баренцевом море наблюдается в октябре, а выход личинок в апреле-мае.
В Белом море цикл размножения изменяется: размножающиеся самки с зародышами на ранних стадиях развития добывались в Кандалакшском заливе в июне, а в Онежском и Двинском - в сентябре.
Инкубация яиц в Белом море продолжается, по-видимому, от 9 до 11 месяцев и по сравнению с другими участками ареала протекает в наибольшем температурном диапазоне (табл. 70). Амплитуда колебаний температуры за период развития эмбрионов достигает в Белом море 13, в прибрежных водах Восточного Мурмана 8.3, в норвежских фиордах 2.2 и в Ирландском море 3°.
| Место наблюдений | I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | Температуры воды, при которой протекает инкубация |
Автор |
| Ирландское море, район Мильпорта | + | + | + | + | * | * | * | * | * | * | * | + | 8-11° | Lebour, 1937а |
| Фиорды Норвегии | + | + | + | + | * | * | + | + | + | + | + | + | -1.1-1.1 | Grieg, 1927 |
| Прибрежные воды Восточного Мурмана | + | + | + | + | * | * | * | * | * | + | + | + | -1.3-7.0 | Наши наблюдения |
| Белое море, Бассейн и заливы | + | + | + | + | + | * | + | + | + | + | + | + | 1.4-11.6 | Наши наблюдения |
Примечание. + время инкубации.
Беломорские размножающиеся самки отличаются от баренцевоморских наиболее крупными размерами тела и наиболее высокой плодовитостью (табл. 71, 72). Относительная плодовитость беломорских самок также более высокая по сравнению с самками большинства баренцевоморских популяций: в Баренцевом море на глубинах более 100 м - 238, на глубинах 50-100 м - 294, на глубинах менее 50 м - 372, в Бассейне же и заливах Белого моря 311 яиц.
| Средняя длина тела (в мм) |
Баренцево море | Бассейн и заливы Белого моря |
||
| глубины более 100 м | глубины 50-100 м | глубины до 50 м | ||
| 47.5 | - | - | - | 735 |
| 42.5 | - | - | - | 421 |
| 37.5 | 353 | - | 360 | 611 |
| 32.5 | 191 | 233 | 290 | - |
| 27.5 | 140 | 149 | 207 | - |
| 22.5 | 80 | 110 | 128 | - |
| 17.5 | 60 | - | 87 | - |
Таким образом, биологическими особенностями беломорских представителей S. spinus служат: некоторое сокращение ареала, появление размножающихся самок летом, более крупные размеры и более высокая индивидуальная плодовитость самок; относительная плодовитость беломорских самок уступает лишь относительной плодовитости самок прибрежной популяции Баренцева моря.
Spirontocaris turgidа (Кr.) являемся типичным обитателем прибрежного мелководья и на Восточном Мурмане образует иногда незначительные поселения среди водорослей на глубинах до 10 м. В Белом море он встречается преимущественно в Кандалакшском заливе на глубинах до 20, а отдельные особи попадаются на глубинах до 51 м (Дерюгин, 1928). В Онежском заливе и в восточной части Белого моря в наших сборах не обнаружен вовсе (рис. 58).
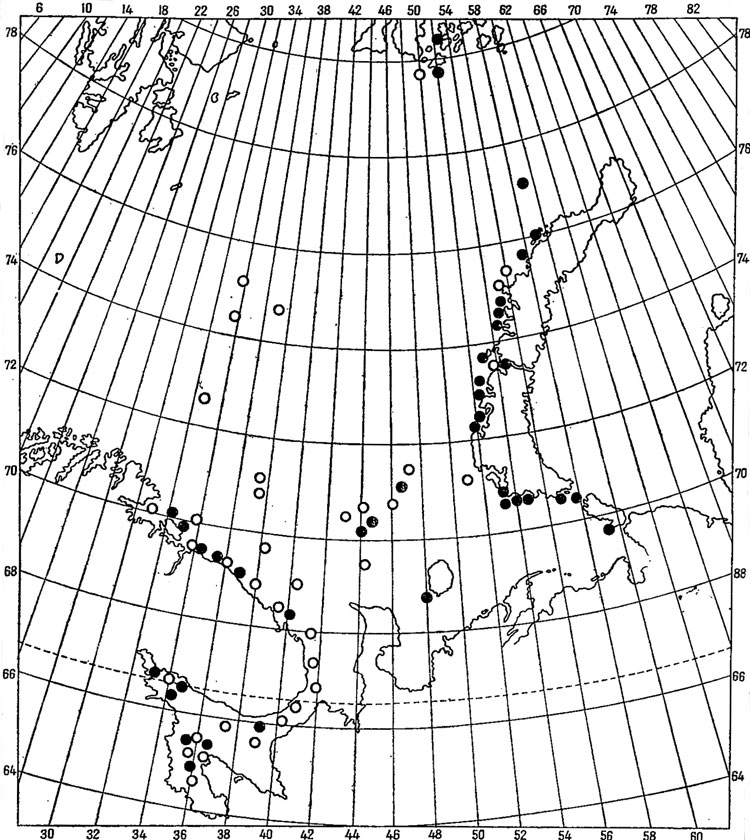
Рис. 58. Распространение Spirontocaris spinus (о) и S. turgida (●) в Баренцевом и Белом морях
Половая зрелость самок в обоих морях наступает на втором году жизни; самцы же, возможно, созревают в возрасте менее 1 года.
В прибрежных водах Восточного Мурмана икрометание происходит чаще всего в октябре, но в некоторые годы оно начинается уже в августе; развитие эмбрионов заканчивается в апреле. В Белом море размножение протекает в общем в те же сроки. На Восточном Мурмане основная масса самок погибает вскоре после первого размножения, т. е. живет около 2 лет, единичные самки размножаются вторично и живут до 3 лет. В некоторые годы продолжительность жизни увеличивается до 4 лет. В Белом море, по-видимому, все самки размножаются только один раз и живут не более 2 лет.
Впервые размножающиеся самки в Баренцевом море имеют длину тела от 16 до 28 мм и вынашивают в среднем (в разные годы) от 91 до 189 зародышей. Вторично размножающиеся самки (на третьем году жизни) имеют длину тела от 24 до 34 мм и вынашивают в среднем (в разные годы) от 198 до 395 зародышей. Самки, размножающиеся в третий раз (на четвертом году жизни), достигают длины 43 мм и вынашивают в среднем 480 зародышей. В Белом море пока найдены только впервые размножающиеся самки (на втором году жизни) с длиной тела от 16 до 27 мм и вынашивающие в среднем по 116 зародышей.
Интересно, что несмотря на удлинение жизни особей этого вида на Мурмане и достижение ими там значительно более крупных размеров,, в среднем беломорские особи оказываются практически такого же размера и даже несколько более крупными. Вообще же в своих средних показателях как по размерам тела, так и по величине плодовитости беломорские и мурманские размножающиеся самки почти не отличаются друг от друга (табл. 72).
| Место наблюдений | n | Длина тела (в мм) | Плодовитость | Корелляция между длиной тела и плодовитостью (r±mr) |
||||
| M±m | σ±mσ | V±mv | M±m | σ±mσ | V±mv | |||
| Spirontacaris spinus | ||||||||
| Прибрежные воды Восточного Мурмана Глубины до 50 м |
116 | 26.0±0.1 | 1.1±0.1 | 4.2±0.3 | 182±7 | 77±7 | 42.4±3.2 | +0.61±0.06 |
| Глубины от 50 до 100 м | 80 | 26.6±0.3 | 3.1±0.2 | 11.7±0.9 | 147±8 | 75±6 | 51.0±4.9 | +0.45±0.09 |
| Глубины более 100 м | 53 | 28.4±0.6 | 4.5±0.4 | 15.8±1.6 | 155±12 | 86±8 | 55.5±6.8 | +0.65±0.08 |
| Бассейн и Кандалакшский залив Белого моря Глубины 12-70 м |
35 | 41.0±1.0 | 6.2±0.7 | 15.1±1.8 | 481±32 | 194±23 | 40.3±5.5 | +0.74±0.08 |
| Spirontacaris turgida | ||||||||
| Прибрежные воды Восточного Мурмана Глубины 0-27 м |
97 | 23.3±0.4 | 3.8±0.3 | 16.3±1.2 | 132±8 | 82±6 | 62.1±6.0 | +0.67±0.06 |
| Кандалакшский залив Белого моря Глубины 2-24 м |
37 | 24.2±0.5 | 3.2±0.4 | 13.2±1.6 | 116±8 | 45±5 | 38.8±5.2 | +0.75±0.07 |
Таким образом, в качестве биологической особенности беломорских S. turgida мы может указать лишь на сокращение экологического ареала, уменьшение численности и сокращение длительности жизни с 3-4 до 2 лет.
Нetairus polaris (Sabine) принадлежит к числу арктических видов и в Баренцевом море распространен чрезвычайно широко: массовые поселения его могут быть обнаружены от нижнего отдела литорали до самых больших глубин (рис. 59). В Кольском заливе он живет на глубине от 5.5 до 274 м (Дерюгин, 1915), у берегов Норвегии от 5 до 775 м и у западной Гренландии - от 0 до 930 м (Nordgaard, 1912; Stephensen, 1913; Dons, 1915; Grieg, 1927, 1935). В водах западной Гренландии самые крупные особи на глубинах от 0 до 290 м имеют длину тела до 60, а на глубинах от 410 до 930 м до 92 мм.
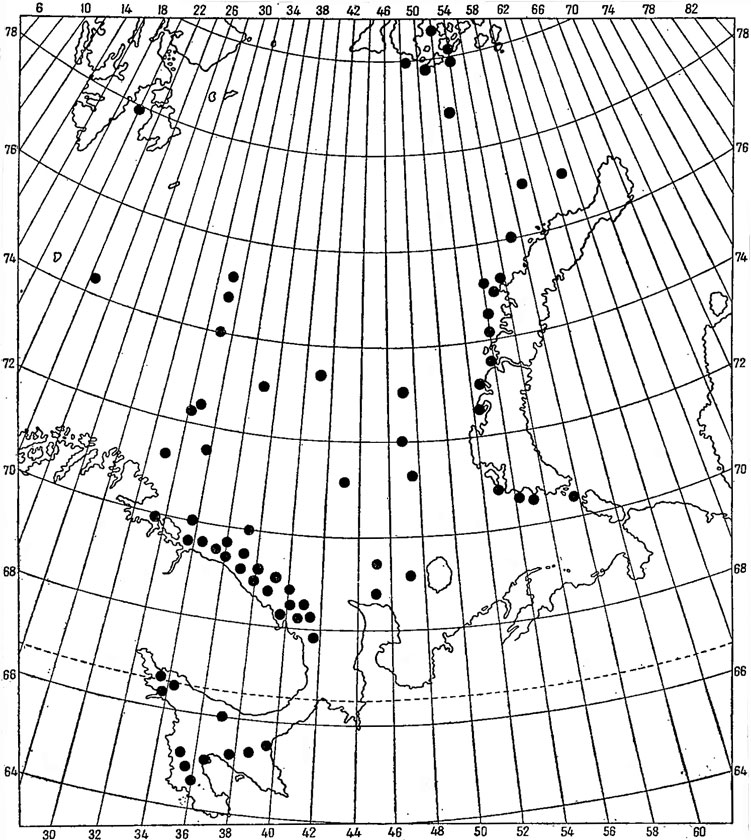
Рис. 59. Распространение Hetairus polaris в Баренцевом и Белом морях
В Баренцевом море самые крупные особи имели длину тела 49, а в Белом море до 55 мм.
В Баренцевом море половая зрелость наступает в возрасте - около 1 года. Самки в большинстве своем размножаются только один раз в жизни и живут около 2 лет. Значительная часть самцов погибает вскоре после первого размножения и живет, таким образом, всего около 1 года. Лишь некоторая часть самцов превращается в самок и участвует в размножении на следующий год. Эти "вторичные" самки имеют продолжительность жизни до 3 лет. В Белом море "вторичных" самок, по-видимому, не бывает, а длительность жизни "первичных" самок в различных районах колеблется от 2 до 4 лет и более. Впрочем, материал по Белому морю был у нас пока весьма ограничен, и высказанные соображения в дальнейшем должны быть уточнены.
В прибрежных водах Восточного Мурмана икрометание замечено в июле и развитие отложенных яиц заканчивается лишь в мае будущего года, т. е. продолжается около 9 месяцев. Средняя длина тела размножающихся самок в различных местообитаниях колеблется от 32.4 до 37.5 мм, а индивидуальная плодовитость от 49 до 110 зародышей. В Белом море наибольшее количество размножающихся самок добыто в августе-сентябре, средняя длина тела этих самок оказалась равной 44.8 мм, а плодовитость в среднем 107 зародышей (табл. 73). Самая мелкая размножающая самка в прибрежных водах Восточного Мурмана имеет длину тела 20 мм, а в Белом море 37 мм.
| Место наблюдений | n | Длина тела (в мм) | Плодовитость | Корелляция между длиной тела и плодовитостью (r±mr) |
||||
| M±m | σ±mσ | V±mv | M±m | σ±mσ | V±mv | |||
| Прибрежные воды Восточного Мурмана Глубины до 50 м |
151 | 34.0±0.3 | 3.1±0.2 | 9.1±5 | 83±2 | 27±2 | 32.5±2.1 | +0.52±0.06 |
| Глубины от 50 до 100 м | 109 | 32.4±0.4 | 3.8±0.3 | 11.7±0.4 | 68±2 | 26±2 | 37.9±2.9 | +0.67±0.05 |
| Глубины более 100 м | 62 | 32.4±0.4 | 3.6±0.3 | 11.0±1.0 | 49±3 | 20±2 | 40.8±4.2 | +0.65±0.07 |
| Район м. Святой Нос Глубины 85-112 м |
18 | 37.5±2.1 | 8.7±1.4 | 23.2±4.1 | 110±11 | 48±8 | 43.6±8.6 | +0.63±0.13 |
| Бассейн и заливы Белого моря Глубины 0-27 м |
12 | 44.8 | - | - | 107 | - | - | - |
Характерным для Hetairus polaris в Белом море являются сокращение численности, ускорение роста и увеличение размеров тела и плодовитости самок. Возможно также, что в южных участках моря (Онежский залив) длительность жизни с 3 лет в прибрежных водах Восточного Мурмана сокращается до 2 лет, а в Бассейне и Кандалакшском заливе, наоборот, длительность жизни увеличивается до 4 и более Лет. Основанием для такого Предположения служит то, что на одном экземпляре был обнаружен прикрепившийся к карапаксу двустворчатый моллюск Anomia squamula L. с ясно различными годовыми кольцами, свидетельствующими о его возрасте 3 лет.
Eualusgaimardi (M. Edw.) образует постоянные и массовые поселения у берегов Восточного Мурмана среди зарослей десмарестии, ламинарий и красных водорослей на глубинах от 4 до 25 м; не менее обычен он и на некотором удалении от берегов, но глубже 120 м почти не встречается. В районе мыса Святой Нос полоса поселений этого вида раздваивается: одна ветвь идет в Белое море, а другая - к побережью Новой Земли, пересекая Канинско-Колгуевское мелководье. На Канинско-Колгуевском мелководье и в прилегающих к нему участках Печорского района типичная форма Е. gaimardi gaimardi смешивается с формой Е. gaimardi gibba (Kroyer), а в прибрежных водах Новой Земли господствующее положение занимает форма Е. gaimardibelcheri (Bell) (рис. 60).
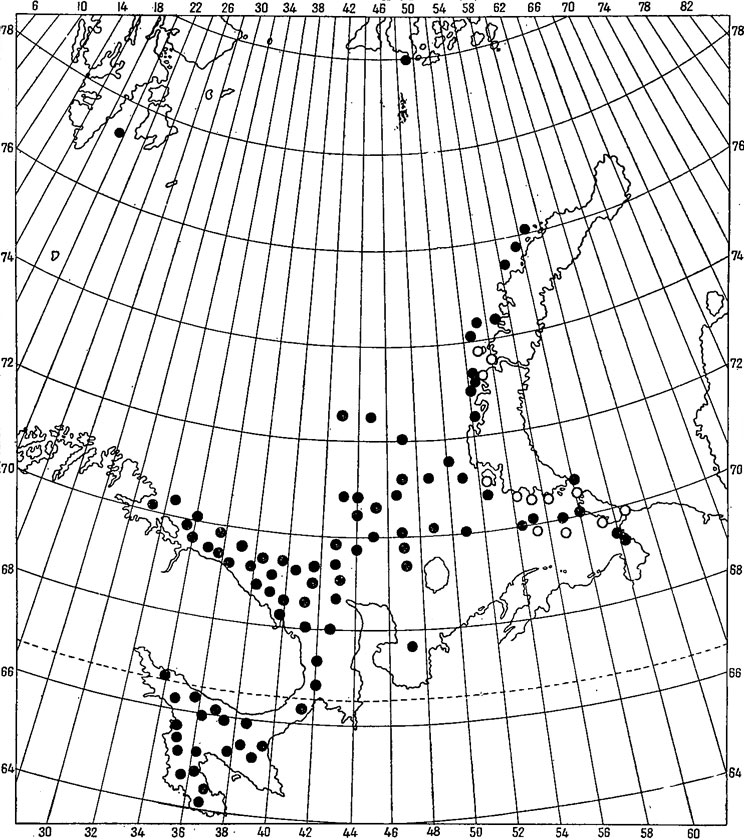
Рис. 60. Распространение Eualus gaimardi (●) и Е. gaimardi belcheri (○) в Баренцевом и Белом морях
Существует представление, что в Белом море имеется только типичная форма этого вида (Бируля, 1910, Алпатов, 1923), но полученный и изученный нами массовый материал противоречит ему. В действительности в Белом море наблюдается на первый взгляд весьма странное положение: обитающие в нем самки принадлежат действительно только к типичной форме, а самцы могут быть всех трех форм. Впрочем, явление это легко объясняется особенностями жизненного цикла, о чем будет сказано несколько позднее.
Поселения Е. gaimardi в Белом море встречаются повсеместно на глубинах от 0 до 140 и даже 160 м, однако наибольшие скопления образуются осенью на глубине от 10 до 50 м.
О сравнительной плотности поселений этого вида можно судить по количеству особей, добываемых за одно траление. В прибрежных водах Восточного Мурмана на глубинах до 50 м за одно траление в среднем добывалось 14 особей (максимально 41), на глубинах от 50 до 100 м - 11 (максимально 33) и на глубинах более 100 м за одно траление добывалось не более одной особи. В Белом море за одно траление в среднем добывалось 26 особей (максимально 141). Очевидно, что в Белом море плотность поселений особей этого вида более высокая, чем в Баренцевом.
Половая зрелость самок наступает в начале второго года жизни и после первого же размножения основная масса их погибает, т. е. продолжительность их жизни не превышает 2 лет. В отдельных случаях единичные самки остаются живыми, размножаются вторично и имеют продолжительность жизни до 3 лет. Самцы становятся половозрелыми на первом году жизни, на втором году жизни часть их превращается в самок, а остальные погибают. Наиболее крупные самки типичной формы имеют длину тела 41.3, а самцы - не более 25 мм. У формы Е. gaimardi belcheri, обитающей в условиях постоянных или почти постоянных отрицательных температур, жизненный цикл значительно удлиняется, что и служит причиной появления у нее некоторых морфологических образований и, в частности, выступа на третьем абдоминальном сегменте. Дело в том, что этот выступ образуется у самцов на втором году жизни и достигает своего полного развития не ранее 3-4 года жизни. В том случае, если превращение самца во "вторичную" самку происходит не на первом, а на втором году жизни, то она получает признаки формы "gibba", а если такое превращение самцов осуществляется в еще более старшем возрасте, то образовавшаяся "вторичная" самка имеет уже все признаки формы "belcheri". В Белом море вторичных самок по еще непонятным причинам не образуется, а длительность жизни самцов увеличивается до 3-4, а возможно и более лет, в результате чего они и успевают развить у себя признаки формы "belcheri", тогда как беломорские самки их никогда не имеют. Таким образом, названные 3 формы Е. gaimardi есть не что иное, как возрастные стадии одной и той же, весьма устойчивой в своих морфологических признаках формы.
Возраст наступления половой зрелости беломорских особей такой же, как и у мурманских, т. е. немногим более 1 года, но продолжительность жизни во многих случаях заметно увеличивается и достигает не менее 4-5 лет. Самые мелкие икряные самки в прибрежных водах Восточного Мурмана имеют длину тела 19.6 мм, а в Белом море 31 мм, что может свидетельствовать об ускорении роста беломорских особей.
Следовательно, более крупные размеры беломорских особей обусловлены повышенной скоростью их роста.
В южной части Баренцева моря икрометание у особей прибрежной популяции происходит в октябре, а в открытом море оно может наблюдаться и в июне. Развитие зародышей заканчивается в апреле-мае. Личинки держатся в планктоне до июля, но молодь появляется уже в июне. В Белом море массовое икрометание замечено в июне-июле и в сентябре-октябре; выход личинок здесь происходит ранней весной.
Беломорские размножающиеся самки отличаются от самок прибрежных вод Восточного Мурмана значительно более крупными размерами и резко возрастающей плодовитостью (табл. 74). Заметим, что увеличение размеров тела беломорских особей обусловлено не только ускорением роста, но и увеличением длительности жизни. По-видимому, правильнее всего считать, что скорость роста более высокая здесь, чем в прибрежных водах Восточного Мурмана, только в первый год жизни, а в последующие годы она сильно замедляется. Размеры тела и плодовитость беломорских самок весьма близки с Е. gaimardi belcheri, населяющей высокоарктическую область и отличающейся замедленным ростом и высокой длительностью жизни.
| Место наблюдений | n | Длина тела (в мм) | Плодовитость | Корелляция между длиной тела и плодовитостью (r±mr) |
Количество зародышей на 1 г живого веса |
||||
| M±m | σ±mσ | V±mv | M±m | σ±mσ | V±mv | ||||
| Прибрежные воды Восточного Мурмана Глубины до 50 м |
144 | 32.5±0.3 | 3.6±0.2 | 11.1±0.7 | 309±18 | 190±13 | 65.6±5.3 | ±0.75±0.04 | 542 |
| Глубины более 50 м | 50 | 31.9±0.3 | 2.1±0.2 | 6.6±0.7 | 316±11 | 70±8 | 22.2±2.6 | ±0.76±0.07 | 464 |
| Район м. Святой Нос Глубины 85-112 м |
137 | 30.6±0.3 | 3.5±0.2 | 11.4±0.8 | 192±9 | 94±7 | 49.0±4.1 | +0.65±0.06 | 362 |
| Канинско-Печорский район, прибрежные воды Новой Земли и другие Сибирские моря (E. Gaimardi belcheri) Глубины 23-191 м |
93 | 44.0±0.3 | 3.3±0.2 | 7.5±0.6 | 486±14 | 132±10 | 27.2±2.1 | +0.72±0.05 | 352 |
| Бассейн и Кандалакшский залив Белого моря Глубины 10-160 м |
119 | 41.1±0.6 | 6.3±0.4 | 15.3±1.0 | 449±15 | 166±11 | 37.0±2.7 | +0.72±0.04 | 368 |
Таким образом, в качестве биологических особенностей беломорских представителей Е. gaimardi можно указать следующее: увеличение численности и экологического ареала, ускорение роста по крайней мере на первом году жизни, увеличение размеров тела и плодовитости, отсутствие "вторичных" самок и увеличение длительности жизни самцов и самок. Неоднократное размножение в течение жизни приводит к тому, что размножающиеся самки начинают появляться здесь не осенью, как это наблюдается на Восточном Мурмане, а в июле и даже в июне. Осенью откладывают яйца только впервые размножающиеся самки. Инкубация зародышей тем не менее заканчивается весной, одновременно как у первично, так и у вторично размножающихся самок.
Сrangon crangon (L.) в Баренцевом море постоянных поселений не образует и встречается лишь единичными особями в прибрежных водах Восточного Мурмана. В Белом море, напротив, этот вид чрезвычайно широко распространен и образует массовые поселения в дельте Северной Двины, в Сухом море, вдоль Летнего берега и в южной части Онежского залива (Паленичко, 1943), имеется он и в Кандалакшском заливе и в южной части Мезенского залива, Всюду он предпочитает песчанистые грунты и занимает глубины от 0 до 54 м. В подобных же условиях С. crangon образует свои массовые поселения у берегов Западной Европы (Wollebaek, 1908; Havinga, 1930; Lebour, 1931). Другой вид этого рода - Сrangоп allmanni Kinahan также населяет прибрежные мелководья, но встречается преимущественно на песчанистых и ракушечных грунтах Воронки и северной части Горла Белого моря; изредка его можно встретить и в прилегающих частях Баренцева моря. В Онежском заливе С. allmanni иногда попадается совместно с С. crangon (рис. 61).
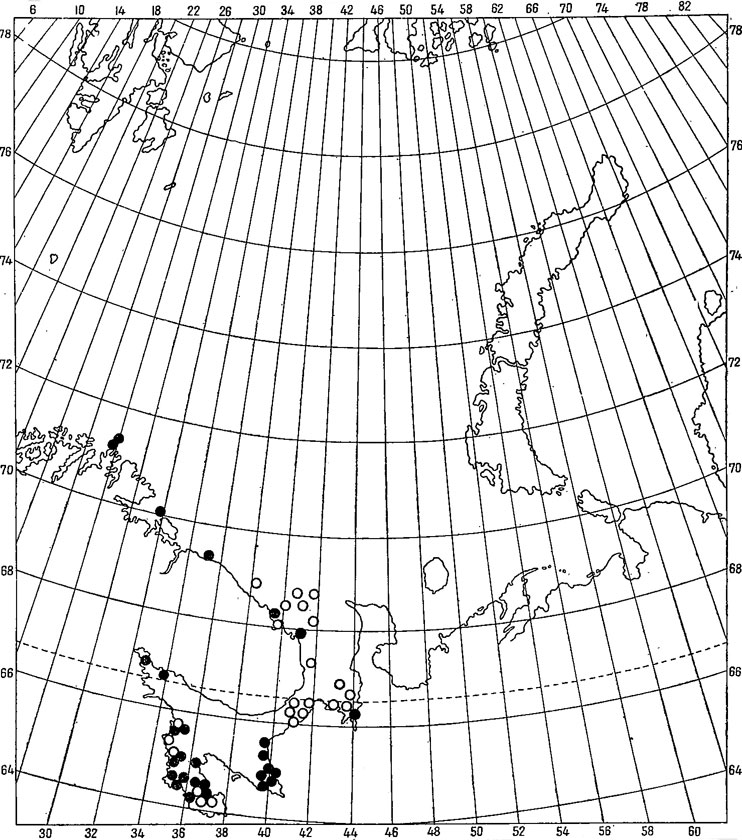
Рис. 61. Распространение Crangon crangon (●) и С. allmanni (○) в Баренцевой и Белом морях
У берегов Западной Европы продолжительность жизни особей С. crangon лишь в редких случаях достигает 4 лет, обычно они не живут более 3 лет. В возрасте около 1 года самки имеют длину 31, 2 лет 55 и 3 лет 75 мм. Самые крупные самки достигают длины 91 мм. Половая зрелость наступает в возрасте около 1 года, а массовая откладка яиц наблюдается в ноябре, в апреле-мае и в июне. Икряные самки встречаются в течение круглого года. При длине тела 40 мм самка вынашивает в среднем 1560 яиц, 50 мм 3120, 60 мм 6240, 65 мм 8820, 70 мм 11500 и 75 мм - 14200 (Havinga, 1930).
В Онежском заливе Белого моря продолжительность жизни особей С. crangon обычно не превышает 1 года, и наиболее крупные из них имеют длину тела до 61 мм (в возрасте 5-6 месяцев). Половая зрелость наступает здесь в возрасте 5-6 месяцев, а откладывание яиц происходит только один раз в году, в октябре. Развитие эмбрионов заканчивается ранней весной, после чего самки погибают и уступают свое место молоди. Длина тела икряных самок колеблется от 46 до 61 мм и каждая самка в среднем вынашивает 2810 зародышей.
Таким образом, беломорские особи С. crangon отличаются от североморских сокращением продолжительности жизни с 3-4 лет до 1 года, ускорением роста и полового созревания и уменьшением индивидуальной плодовитости. Икрометание и развитие эмбрионов в Северном море протекают в любое время года, а в Белом море икрометание наблюдалось только в октябре, а развитие эмбрионов идет в самое холодное время года, с октября по май, т. е. в течение 7-8 месяцев. Заметим, что эмбрионы североморских самок при температуре 5-6° заканчивают свое развитие не более чем в 13 недель, а при 16° за 4.5 недели (Havinga, 1930).
По сравнению со всеми другими участками довольно обширного ареала инкубация яиц в Белом море протекает при самых низких температурах воды (табл. 75), чем и объясняется ее большая продолжительность.
| Место наблюдений | Месяцы | Температуры воды, при которой протекает инкубация |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Черное море, район Севастополя | * | * | + | + | + | * | * | * | * | * | * | * | 6.6-13.5° | Зернов, 1913 |
| Средиземное море, Неаполитанский залив | + | + | + | * | * | * | * | * | * | * | * | * | * | Lo Bianco, 1909; Meyer, 1934, 1935 |
| Прибрежные воды Франции, Бельгии и Голландии* | + | + | + | + | + | + | + | + | + | + | + | + | 5-20 | Havinga, 1930; Lloyd a. Jonge, 1947 |
| Район Гельголанда** | * | * | * | + | + | + | + | + | + | + | + | * | 5-19 | Ehrenbaum, 1890 |
| Прибрежные воды Дании, Зунд | * | * | * | * | + | * | + | + | * | * | * | * | 6.9-17.9 | Thorson, 1946 |
| Фиорды Норвегии, район Бергена | * | ? | + | + | + | * | * | * | * | * | * | * | 4-8 (?) | Grieg, 1927 |
| Белое море, Онежский залив*** | + | + | + | + | + | * | * | * | * | + | + | + | -1.4+6.0 | Наши наблюдения |
Примечание. + время инкубации.
* (В течение года 3 периода массового размножения.)
** (В течение года 2 периода массового размножения.)
*** (В течение года только 1 период размножения.)
Sсlerocrangon boreas (Phipps) служит представителем арктической фауны и довольно широко распространен в прибрежных водах Восточного Мурмана; изредка он встречается и в открытом море, но, по-видимому, далее 72° с. ш. не идет. Может жить на глубинах от 0 до 180 м, но постоянные поселения образует только на глубинах 15-20 м, предпочитая устьевые участки губ, где за одно траление может быть добыто до 56 особей. В среднем в Баренцевом море за одно траление в местообитаниях этого вида добывалось 9 особей.
В Белом море S. boreas более многочислен и образует массовые поселения в восточной и северо-восточной частях Бассейна на глубинах 35-40 м и в северной части Онежского залива на глубинах 20-35 м. Единичные крупные особи встречаются повсеместно и чаще всего в Кандалакшском заливе на глубинах до 220 м. В среднем за одно траление в местообитаниях этого вида в Белом море добывалось 21, а в северо-восточной части Бассейна 113 особей.
Другой вид этого рода - Sclerocrangen ferox (Sars) населяет преимущественно северо-восточную часть Баренцева моря и в Белом море был обнаружен лишь один раз (рис. 62), поэтому о нем здесь говорить не будем.
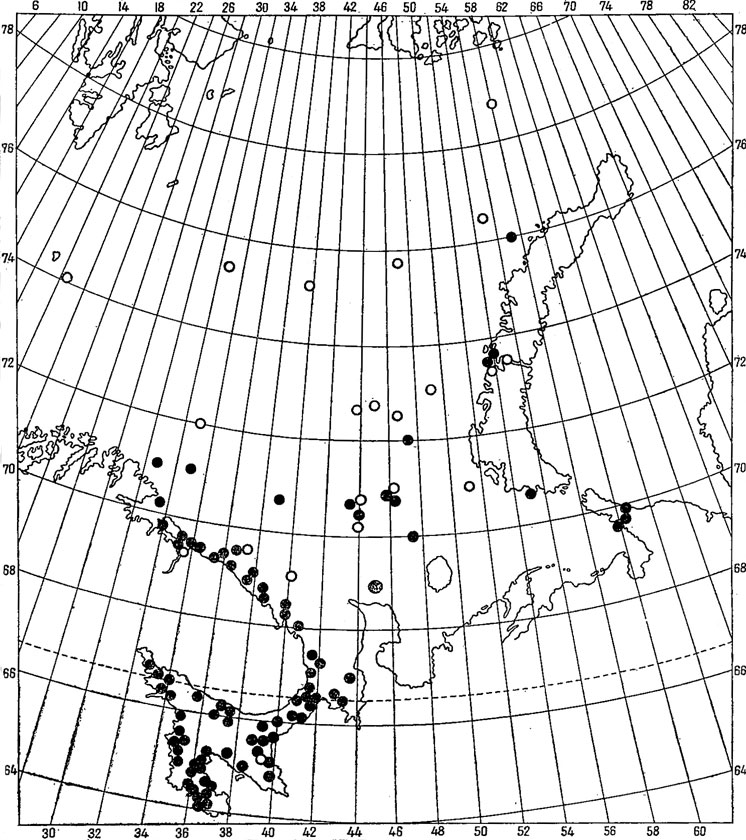
Рис. 62. Распространение Sclerocrangon boreas (●) и S. ferox (о) в Баренцевом и Белом морях
Половая зрелость самок S. boreas наступает в возрасте не менее 2 лет; по достижении половой зрелости самки размножаются 1 или 2 раза и живут не более 4 лет. Самые крупные самки в Баренцевом море имеют длину тела 86, а в Белом 80 мм.
Массовая откладка яиц в прибрежных водах Восточного Мурмана происходит в декабре-январе, и развитие эмбрионов заканчивается в мае - июне, т. е. продолжается 4-5 месяцев. В Белом море икрометание наблюдалось в августе-сентябре и позднее, развитие зародышей заканчивалось ранней весной, т. е. продолжалось около 9-10 месяцев. Как в Баренцевом, так и в Белом морях икрометание может происходить и в другое время года.
Икряные самки в Баренцевом море имели длину тела от 70 до 86 мм и вынашивали в среднем 195 зародышей; беломорские самки имели длину тела от 69 до 80 мм и вынашивали в среднем 161 зародыша. Средняя длина тела баренцевоморских размножающихся самок составляет 69.5 мм, а беломорских - 76.5 мм.
Таким образом, в отношении беломорских S. boreas можно с уверенностью сказать, что от баренцевоморских они отличаются более высокой численностью и более крупными средними размерами тела размножающихся самок.
Судя по коллекциям Зоологического института АН ССCP, указанные здесь размеры тела баренцевоморских особей, по-видимому, не являются предельными. Во всяком случае, в конце прошлого и в начале текущего столетия в прибрежных водах Мурмана встречались особи с длиной тела до 106.2 мм. Икряные самки, добытые в районе мыса Святой Нос на глубинах 35-87 м, имели длину тела до 90.5-98.5 мм и вынашивали от 163 до 314 зародышей. Возможно, что столь крупные размеры особей были обусловлены наступлением половой зрелости в более старшем возрасте и вообще удлинением жизни под влиянием имевшего место периода похолодания.
Sabinea septemcarinata (Sab.) подобно предшествующему виду служит представителем арктической фауны. В Баренцевом море она широко распространена и может быть встречена всюду и на любой глубине. Наиболее частые и массовые местонахождения прижимаются к побережью Восточного Мурмана и затем по краю Канинско-Колгуевского мелководья идут на северо-восток. В Белом море поселения этого вида также широко распространены, за исключением Горла и южной части Онежского залива, где нам не встречались даже единичные особи.
Другой вид этого рода - Sаbinea sarsi Smith, в незначительном количестве обитает в восточной части Баренцева моря, а в Белом море до сих пор не обнаружен (рис. 63).
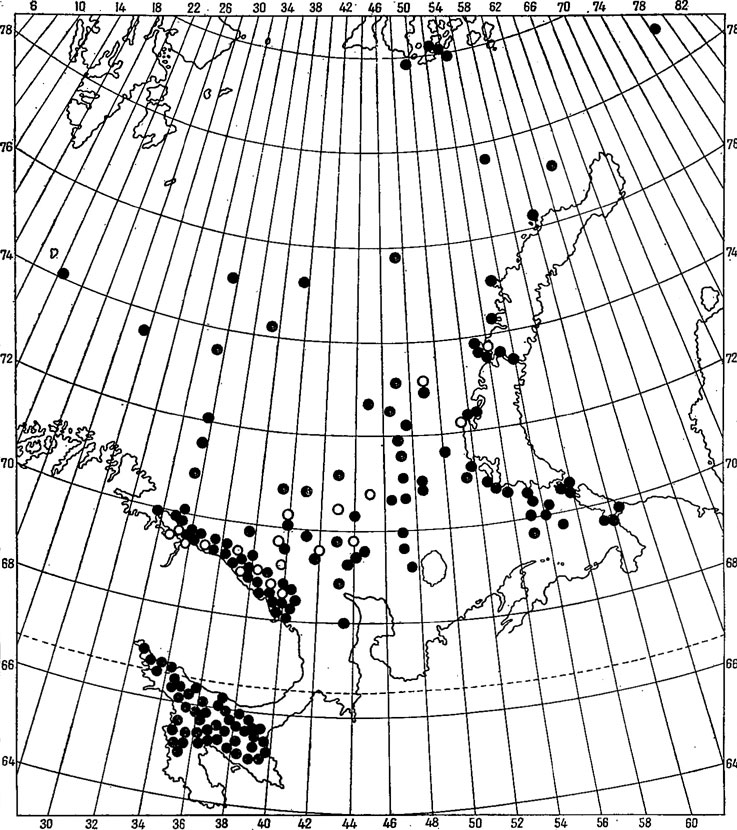
Рис. 63. Распространение Sabinea septemcarinata (●) и S. sarsi (○) в Баренцевом и Белом морях
В прибрежных водах Восточного Мурмана на глубинах более 100 м за одно траление в местообитаниях S. septemcarinata добывалось в среднем 5 (максимально 54), на глубинах 50-100 м 10 (максимально 65) и на глубинах до 50 м 4 особи (максимально 12). По-видимому, оптимальными для этого вида можно считать глубины 50-100 м.
В Белом море особенно массовые поселения этой креветки имеются в восточной части Бассейна на глубинах 40-60 м; за одно траление здесь добывалось до 121 особи, в среднем каждый трал в местах поселений этого вида приносил 12 особей.
Самые крупные особи в Баренцевом море имеют длину тела 81, а в Белом море 85 мм. Продолжительность жизни в обоих морях в общем одинакова и обычно не превышает 3 лет, хотя отдельные особи, очевидно, могут жить до 4 лет и более. До двухлетнего возраста беломорские особи крупнее баренцевоморских на 10, а в возрасте 3 лет только на 4 мм.
Половая зрелость обычно наступает в начале второго года жизни, и в течение жизни самки размножаются 2 и в некоторых случаях 3 раза - по одному разу в год. В Баренцевом море икрометание происходит с мая по сентябрь, а развитие отложенной икры заканчивается к июню-июлю будущего года, т. е. продолжается около 12 месяцев. В Белом море икрометание происходит с апреля по сентябрь и развитие эмбрионов также продолжается около 12 месяцев.
В различных местообитаниях прибрежных вод Восточного Мурмана средняя длина тела размножающихся самок колеблется от 49.9 до 52.3 мм, а плодовитость - от 182 до 193. В Белом море размножающиеся самки значительно более крупные: средняя длина их составляет 65.7 мм и они вынашивают в среднем по 341 зародыша (табл. 76), т. е. почти вдвое больше, чем на Мурмане. Самые мелкие размножающиеся самки в прибрежных водах Восточного Мурмана имеют длину тела 36 мм, а в Белом море 55 мм.
| Место наблюдений | n | Длина тела (в мм) | Плодовитость | Корелляция между длиной тела и плодовитостью (r±mr) |
||||
| M±m | σ±mσ | V±mv | M±m | σ±mσ | V±mv | |||
| Прибрежные воды Восточного Мурмана Глубины 50-100 м |
82 | 49.9±0.7 | 5.2±0.5 | 10.4±1.1 | 182±12 | 82±8 | 45.1±5.4 | +0.51±0.10 |
| Глубины более 100 м | 49 | 52.3±0.4 | 3.7±0.3 | 7.1±0.6 | 193±11 | 102±8 | 59.0±6.0 | +0.21±0.11 |
| Бассейн и заливы Белого моря Глубины 20-220 м |
48 | 65.7±0.8 | 5.6±0.6 | 8.5±0.6 | 341±18 | 128±13 | 53.1±6.7 | +0.59±0.09 |
Таким образом, беломорские представители этого вида отличаются от мурманских повышенной численностью, ускоренным ростом, более крупными размерами тела и более высокой плодовитостью.
Pagurus pubesсens (Кr.) принадлежит к числу арктическо-бореальных видов и широко распространен как в Баренцевом, так и в Белом морях. В Южной части Баренцева моря он обитает от нижнего отдела литорали до самых больших глубин, на северо-западе поднимается до Шпицбергена, а вдоль побережья Новой Земли доходит до 77° с. ш. В центральной части моря далее 75° с. ш., по-видимому, не идет (рис. 64).
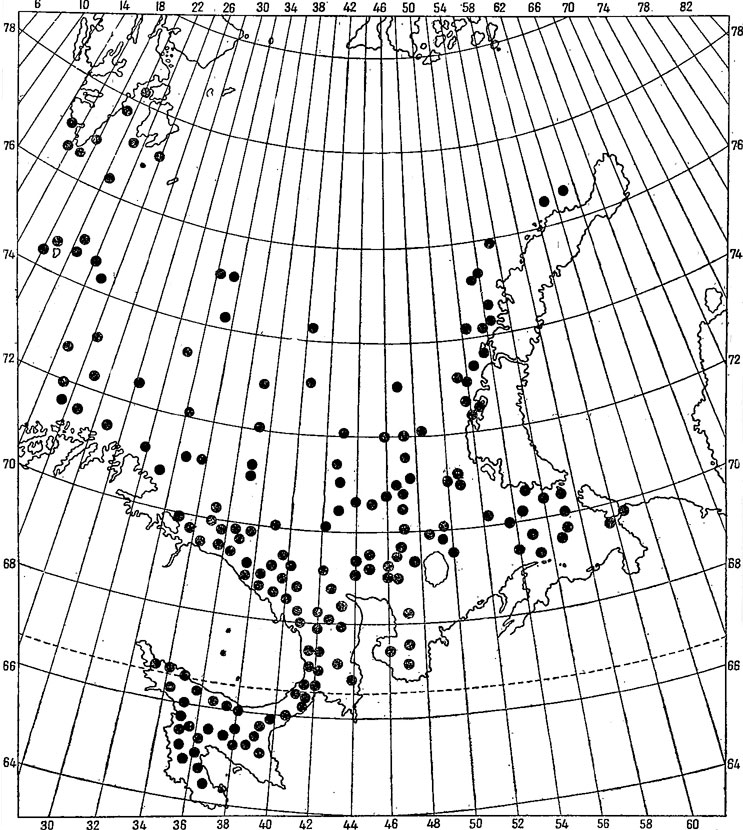
Рис. 64. Распространение Pagurus pubescens в Баренцевом и Белом морях
В Белом море встречается повсюду, но особенно многочисленные скопления образует в северной половине Онежского залива и в примыкающих к ней частях Бассейна на глубинах 29-54 м. Нигде на литорали и на глубинах менее 12 м особи этого вида южнее Горла не найдены. К. М. Дерюгин (1928) подчеркивал также, что глубже 100 м P. pubescens в Белом море не идет, тогда как в Кольском заливе он встречается до глубины 330 м. В Воронке и в северной части Горла Белого моря этот вид может быть встречен в нижнем отделе литорали и на глубинах до 70 м.
В среднем за одно траление в местообитаниях рака-отшельника добывалось в прибрежных водах Восточного Мурмана на глубинах более 100 м 22 (максимально 131) и на глубинах до 50-60 м 12 особей (максимально 138). В Воронке и Горле Белого моря за одно траление в среднем добывалось 66 (максимально 223), а в Бассейне и заливах 19 особей (максимально 71).
В среднем плотность поселений беломорских особей по существу не уступает плотности поселений баренцевоморских особей. Особенно высокая численность этого вида замечена в северной части Горла.
Половая зрелость самок во всех популяциях может наступать в возрасте нескольких месяцев и не позднее начала второго года жизни. От общего количества самок на первом году жизни становятся половозрелыми и размножаются 61-66% в Баренцевом море и 74% в Бассейне и заливах Белого моря. По достижении половой зрелости самки размножаются ежегодно и имеют продолжительность жизни в Баренцевом море, в Бассейне и заливах Белого моря около 3 лет. В Воронке и в северной части Горла особи живут обычно 4-5 лет.
Икрометание в Баренцевом море происходит в октябре - декабре, а в Бассейне и заливах Белого моря - в сентябре-октябре. Развитие эмбрионов заканчивается и личинки выходят в планктон в начале мая.
Продолжительность периода инкубации яиц в различных частях ареала изменяется в полном соответствии с температурой воды, при которой протекает развитие эмбрионов (табл. 77). С момента откладывания яиц и до выхода личинок проходит в Северном и Норвежском морях 7, в прибрежных водах Восточного Мурмана 8 и в Бассейне и заливах Белого моря 9 месяцев.
| Место наблюдений | Месяцы | Температуры воды, при которой протекает инкубация |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Северное море | + | + | + | + | + | * | * | * | * | * | + | + | 2-12 (?) | Schellenberg, 1928 |
| Прибрежные воды северной Норвегии | + | + | + | + | * | * | * | * | * | + | + | + | 1.6-11.5 (средняя многолетняя) | Schellenberg, 1928 |
| Восточный Мурман, район губы Дальне-Зеленецкой |
+ | + | + | + | + | * | * | * | * | + | + | + | -1.0+8.0 | Наши наблюдения |
| Белое море, Бассейн и заливы | + | + | + | + | + | * | * | * | + | + | + | + | -1.4+7.7 | Наши наблюдения |
Примечание. +время инкубации.
В различных местообитаниях прибрежных вод Восточного Мурмана средняя длина карапакса размножающихся самок колеблется от 10.2 до 16.0 мм и каждая самка вынашивает в среднем от 1574 до 3341 зародыша. Размеры тела и плодовитость увеличиваются по мере увеличения глубины расположения местообитания. В Белом море средняя длина карапакса размножающихся самок составляет 14.3 мм, а индивидуальная плодовитость 1878 (табл. 78). Это создает совершенно определенное впечатление, что беломорские самки уступают мурманским как по величине тела, так и по плодовитости. Однако на первом году жизни наблюдается прямо противоположное: беломорские самки оказываются и крупнее и плодовитее мурманских. В этом возрасте беломорские размножающиеся самки в 1.5-2 раза крупнее и в 3-5 раз более плодовиты. На втором году жизни размеры тела и плодовитость мурманских и беломорских размножающихся самок становятся практически одинаковыми. Уменьшение средних размеров тела беломорских размножающихся самок обусловлено их повышенной смертностью после первого же размножения и господством в популяции впервые размножающихся самок, имеющих возраст менее 1 года.
| Место наблюдений | n | Длина карапакса (мм) | Плодовитость | Корелляция между длиной карапакса и плодовитостью (r±mr) |
||||
| M±m | σ±mσ | V±mv | M±m | σ±mσ | V±mv | |||
| Самки всех возрастных групп | ||||||||
| Прибрежные воды Восточного Мурмана Глубины 0-60 м |
99 | 10.2±0.6 | 5.8±0.4 | 56.9±5.2 | 1574±212 | 2116±150 | 140.8±19.1 | +1.00±0 |
| Глубины более 60 м | 84 | 16.0±0.6 | 5.2±0.4 | 32.5±2.8 | 3341±284 | 2560±200 | 76.6±8.9 | +0.92±0.02 |
| Воронка и северная часть Горла Глубины 20-50 м |
4 | 9.6 (?) | - | - | 576 (?) | - | - | - |
| Бассейн и заливы Глубины 29-54 м |
67 | 14.3±0.3 | 2.6±0.2 | 18.2±1.6 | 1878±122 | 1001±86 | 53.3±5.8 | +0.91±0.02 |
| Самки на первом году жизни | ||||||||
| Прибрежные воды Восточного Мурмана Глубины до 60 м |
65 | 6.1±0.2 | 1.6±0.1 | 26.2±2.4 | 286±16 | 132±11 | 46.2±4.8 | +0.64±0.07 |
| Их открытая часть Глубины 60-220 м |
29 | 9.3±0.4 | 2.1±0.3 | 22.6±3.1 | 547±89 | 490±64 | 89.7±6.4 | +0.81±0.06 |
| Бассейн и залив Белого моря Глубины 12-60 м |
48 | 12.9±0.2 | 1.4±0.1 | 10.9±1.1 | 1310±59 | 400±40 | 35.5±3.3 | +0.56±0.10 |
| Самки на втором году жизни | ||||||||
| Прибрежные воды Восточного Мурмана Глубины до 60 м |
23 | 17.7±0.4 | 1.9±0.2 | 10.7±1.4 | 3090±199 | 1112±141 | 32.8±4.4 | -0.07±0.18 |
| Их открытая часть Глубины 60-220 м |
42 | 19.2±0.3 | 1.9±0.2 | 9.9±1.1 | 4246±193 | 1272±137 | 30.0±3.5 | +0.22±0.14 |
| Бассейн и залив Белого моря Глубины 12-60 м |
17 | 18.5±0.3 | 1.1±0.2 | 5.9±1.0 | 3471±88 | 362±62 | 10.4±1.8 | +0.46±0.19 |
Таким образом, биологическими особенностями беломорских представителей рака-отшельника являются: ускорение роста и увеличение плодовитости на первом году жизни и резкое замедление роста в последующие годы.
Нуas araneus (L.) в Норвежском море является настоящим прибрежным обитателем и живет преимущественно на литорали (Appelöf, 1906). К востоку от Рыбачьего полуострова он приобретает некоторые новые морфологические черты, что и позволило А. А. Бируле (1897) выделить всех особей этой восточной популяции в самостоятельный вариетет Н. araneus v. hoeki.
Н. araneus hoeki вдоль берегов Восточного Мурмана образует массовые поселения от нижнего отдела литорали до глубин 8-10 м; единичные особи могут быть встречены и на больших глубинах. На зиму поселения этого краба перемещаются несколько ниже: от границы отлива до глубин 25-30 м. В Воронке и в северной части Горла характер поселений сохраняется в почти неизменном виде: различие состоит в том, что на литорали здесь живет только молодь, а взрослые особи, как правило, живут ниже линии отлива. В Бассейне Белого моря, в Кандалакшском и Двинском заливах массовых поселений нигде не образует, хотя и встречается в сублиторали повсеместно до глубины 50 м. В Онежском заливе сплошные поселения этого вида имеются в районе островов Соловецкого, Б. Жужмуйского, Кузовов, Кондострова, Мягострова и др., доходя до самой южной части залива (остров Шоглы). Обычно эти поселения располагаются на глубинах от 8-12 до 30-35 м, но в некоторых случаях (около островов Соловецкого, Б. Жужмуйского, Русского Кузова и др.) большое количество особей можно обнаружить около самой линии отлива, но всегда ниже ее. На литорали Бассейна и заливов Белого моря Н. araneus hoeki найден не был.
Во всех рассмотренных случаях Н. araneus hoeki всегда поселялся только в прибрежной зоне, а молодь его часто была связана с литоральной зоной. Особое положение занимает район Канинско-Колгуевского мелководья и прилегающая к нему юго-восточная часть Баренцева моря. Здесь, к северу и к северо-востоку от мыса Канин Нос располагаются массовые поселения краба, занимающие обширные площади до глубины 250 м. Каждое траление в этом районе приносит до 100 и более особей.
Другой вид этого рода - Нуas соаrctatus Leach массовые поселения образует к западу от Рыбачьего полуострова, но в незначительном количестве доходит примерно до меридиана мыса Святой Нос. Отдельные особи могут быть встречены и в Белом море (рис. 65). Собранный здесь материал оказался весьма ограниченным, и поэтому оказать что-либо определенное о биологических особенностях беломорских Н. coarctatus пока не представляется возможным.
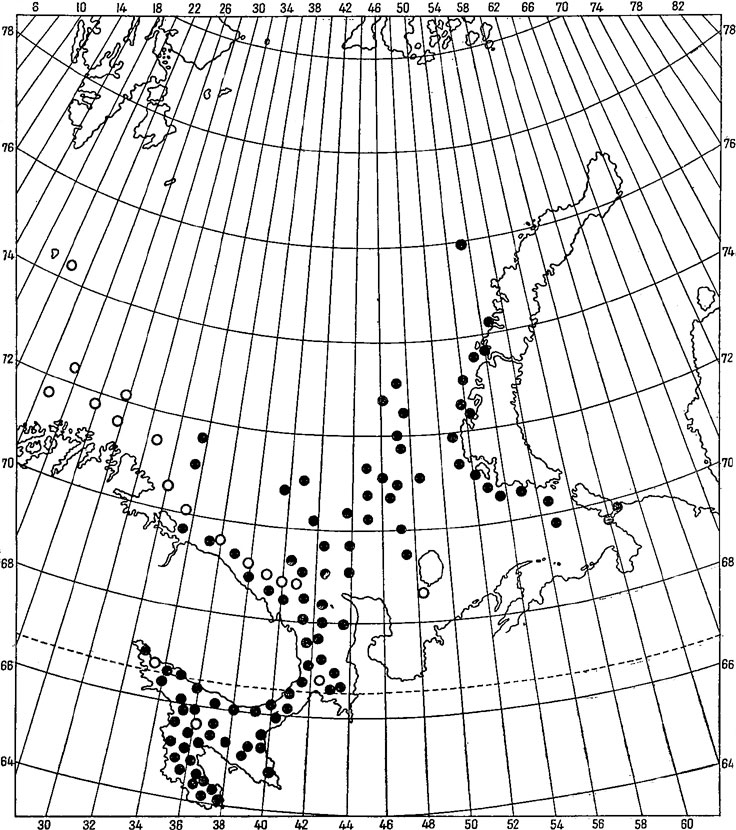
Рис. 65. Распространение Hyas araneus (●) и Н. coarctatus (○) в Баренцевом и Белом морях
Продолжительность жизни особей Н. araneus прибрежной популяции Восточного Мурмана составляет 10-11 лет, при достижении половой зрелости в возрасте 3-4 лет. Возраст наступления половой зрелости и продолжительность жизни особей других популяций пока неизвестны.
Икрометание в Баренцевом и Белом морях происходит практически одновременно, в мае-июне, и развитие отложенной икры заканчивается не ранее мая (иногда в апреле) будущего года, т. е. продолжается до 11 месяцев. Икряные самки всюду встречаются в течение круглого года. В других частях ареала выход личинок наблюдался чаще всего в апреле.
Минимальная температура воды в период выхода личинок в различных частях ареала изменяется весьма существенно и самых низких показателей достигает в Белом море: если у побережья Англии температура воды в это время редко бывает ниже 6-7°, то в Белом море она. едва достигает 0° (табл. 79).
| Место наблюдений | Месяцы | Средние месячные температуры воды, при которых протекает выход личинок |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Ирландское море | * | * | * | + | * | * | * | * | * | * | + | + | 6-9° (?) | Jackson, 1913 |
| Прибрежные воды Шотландии | * | * | * | * | + | * | * | * | * | * | * | * | 7-9 | Jorgengen, 1923; Williamson, 1911 |
| Восточный Мурман, район губы Дальне-Зеленецкой |
+? | +? | + | + | * | * | * | * | * | * | * | * | 3-4 | Dons, 1915; Runnström, 1930 |
| Прибрежные воды Восточного Мурмана, район губы Дальне-Зеленецкой |
* | * | * | * | + | + | * | * | * | * | * | * | 2-3 | Наши наблюдения |
| Белое море. Бассейн и заливы | * | * | * | * | + | + | * | * | * | * | * | * | 0-6 | Наши наблюдения |
Примечание. + время выхода личинок.
Наиболее крупные размеры тела и наиболее высокую плодовитость имеют самки, живущие в непосредственной близости к берегам Восточного Мурмана; на некотором удалении от берегов и на больших глубинах размеры тела и плодовитость самок снижаются (табл. 80). Самые мелкие и наименее плодовитые самки населяют Онежский залив Белого моря. В Кандалакшском заливе, насколько об этом можно судить на основании небольшого количества добытого материала, размножающиеся самки крупнее, чем в Онежском заливе и более, плодовиты: длина карапакса колебалась у них от 58 до 63 мм и каждая из них вынашивала от 6972 до 17369 зародышей. Самые мелкие размножающиеся самки имели длину карапакса в различных местообитаниях Восточного Мурмана от 43 до 49, в Воронке Белого моря 49 и в Онежском заливе 37 мм.
| Место наблюдений | n | Длина карапакса (в мм) | Плодовитость | Корелляция между длиной карапакса и плодовитостью (r±mr) |
||||
| M±m | σ±mσ | V±mv | M±m | σ±mσ | V±mv | |||
| Прибрежные воды Восточного Мурмана Литораль и сублитораль до глубины 40 м |
157 | 78.3±0.7 | 8.2±0.5 | 10.5±0.6 | 41684±1560 | 19500±1112 | 46.8±3.2 | 0.00 |
| Открытая часть. Глубины 70-250 м | 42 | 57.4±1.0 | 6.3±0.7 | 10.2±1.1 | 17498±1115 | 7250±788 | 41.4±5.2 | +0.42±0.14 |
| Белое море. Юго-западная часть Воронки. Глубины 6-8 м |
36 | 64.6±0.9 | 5.8±0.7 | 9.0±1.1 | 21111±1863 | 11180±1315 | 52.5±5.4 | +0.39±0.14 |
| Онежский залив. Глубины 8-35 м | 70 | 48.5±0.7 | 5.6±0.5 | 11.5±1.0 | 12498±702 | 5900±500 | 47.2±4.9 | +0.54±0.08 |
На 1 г живого веса самки в среднем откладывается яиц: в прибрежной популяции Восточного Мурмана 390, в популяции из района Канинской и Гусиной банок 446, в популяции из Воронки Белого моря 320 и в популяции из Онежского залива Белого моря 521. Это может служить свидетельством того, что условия жизни в Онежском заливе угнетающе действуют на рост организма и способствуют деятельности гонады.
Приведенный здесь материал позволяет отметить следующие биологические особенности беломорских представителей этого вида: общее сокращение экологического ареала и отсутствие массовых литоральных поселений, замедление скорости роста, уменьшение размеров тела и индивидуала ной плодовитости, сопровождаемое заметным увеличением относительной плодовитости.
Verruca strömia (О. F. Müll) в Баренцевом море распространена вдоль, всего южного берега к западу от 44 меридиана; севернее 71° с. ш. она на встречается. В пределах столь ограниченной области этот вид может быть встречен от нижнего отдела литорали до глубины 200 м. На литорали и в верхнем отделе сублиторали он поселяется обычно на нижней и боковых сторонах камней; в более низких горизонтах животные прикрепляются уже только к верхней стороне камней, к раковинам моллюсков и к домикам других беспозвоночных.
В кутовых участках губ V. strömia обычно отсутствует и наиболее плотные поселения образует у границы отлива вдоль открытых и полуоткрытых берегов Восточного Мурмана. Подобный же характер вертикального распределения сохраняется в общих чертах и в Воронке и в северной части Горла, с той лишь разницей, что у границы отлива сплошных поселений, как на Мурмане, по-видимому, не бывает. Южнее Горла, в Бассейне и заливах Велого моря, у границы отлива и тем более на литорали V. strömia не найдена. Более или менее постоянные и плотные поселения она образует здесь в сублиторали на глубинах 17-30 м в местах с быстрыми течениями, используя в качестве субстрата мелкие камни, раковины моллюсков и домики других животных.
Максимальная плотность поселений на Восточном Мурмане составляет 390, а в Белом моще 250-280 особей на 100 см2.
В Баренцевом и Белом морях продолжительность жизни особей обычно не превышает 3-4 лет, а половая зрелость наступает в начале второго года жизни.
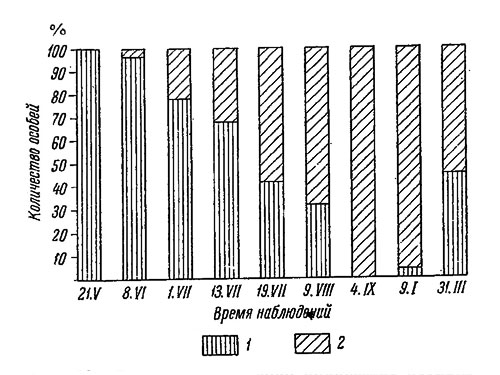
Рис. 66. Сезонные изменения количества размножающихся (1) и неразмножающихся (2) особей (в процентах от общего количества взрослых) Verruca stromia, обитающих у границы отлива на закрытых берегах Восточного Мурмана
Откладывание яиц в мантийную полость в Баренцевом море начинается в самый разгар зимы, в январе (рис. 66). В это время 4% взрослых особей имеют в полости мантии развивающихся эмбрионов, в марте таких особей становится половина, в мае все взрослые особи вынашивают зародышей. Выход науплиусов начинается в июне и заканчивается в августе. Период инкубации яиц, отложенных в январе, продолжается около 5 месяцев, а отложенных в июне - не более 3 месяцев (рис. 67). Массовое прикрепление личинок к субстрату на Восточном Мурмане наблюдалось в течение сентября; в это же время оно происходит и в Онежском заливе Белого моря.
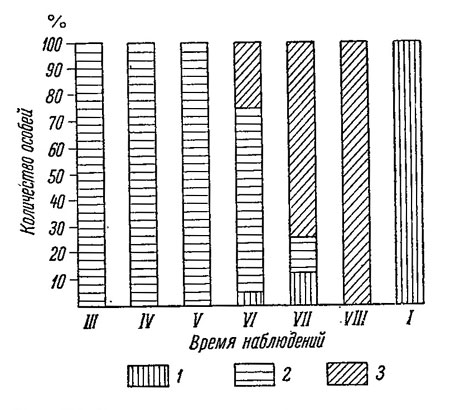
Рис. 67. Сезонные изменения соотношения между особями Verruca stromia (в процентах от общего количества размножающихся), имеющими в выводковых сумках зародыши на различных стадиях развития. 1 - яйца и стадии дробления, 2 - стадии сегментации, 3 - вполне сформировавшиеся науплиусы
Беломорские размножающиеся особи отличаются от баранцевоморских самыми мелкими размерами и самой маленькой плодовитостью (табл. 81).
| Место наблюдений | n | Длина карапакса (в мм) | Плодовитость | Корелляция между длиной карапакса и плодовитостью (r±mr) |
||||
| M±m | σ±mσ | V±mv | M±m | σ±mσ | V±mv | |||
| Баренцево море. К северу от мыса Святой Нос. Глубина 85 м. На раковинах моллюсков |
22 | 5.2±0.2 | ±1.0±0.1 | 19.2±3.0 | 1409±203 | ±954±145 | 67.7±14.1 | ±0.72±0.10 |
| Восточный Мурман. У границы отлива на открытых берегах. На камнях |
79 | 6.3±0.1 | ±1.1±0.1 | 17.5±1.6 | 1351±73 | ±580±46 | 42.9±4.5 | ±0.68±0.06 |
| У границы отлива на закрытых берегах. На камнях |
85 | 5.4±0.1 | ±1.0±0.1 | 18.5±1.5 | 874±48 | ±446±34 | 51.0±4.8 | ±0.51±0.08 |
| Белое море. Онежский залив. К западу от острова Соловецкого. Глубина 30 м. На камнях |
65 | 4.7±0.1 | ±1.0±0.1 | 21.3±1.9 | 766±57 | ±464±40 | 60.5±6.9 | ±0.81±0.04 |
Вalanus balanoides L. является типичным литоральным обитателем и образует более или менее массовые поселения вдоль всех побережий Восточного Мурмана и Белого моря. Отличительной чертой беломорских поселений может служить их сравнительно небольшая плотность и биомасса (табл. 82).
| Уровень над 0 глубины (в см) |
Количество особей (в шт/м2) |
Биомасса (в г/м2) |
Средний вес одной особи (в г) |
| Восточный Мурман. Валунно-каменистые россыпи. Июль 1946 | |||
| 326 | 43 | 17.5 | 0.41 |
| 314 | 22100 | 4432.0 | 0.20 |
| 306 | 31200 | 4770.6 | 0.15 |
| 276 | 33600 | 2757.0 | 0.08 |
| 160 | 27300 | 625.8 | 0.02 |
| 80 | 12560 | 259.6 | 0.02 |
| Белое море. Кандалакшский залив. Каменистые россыпи. Май 1946 | |||
| 210 | 10235 | 1656.4 | 0.16 |
| 200 | 9000 | 2215.5 | 0.25 |
| 190 | 17375 | 831.3 | 0.05 |
| 160 | 13113 | 1155.1 | 0.09 |
| 130 | 14774 | 867.8 | 0.06 |
| 100 | 11100 | 1002.0 | 0.09 |
| 70 | 27639 | 574.9 | 0.02 |
На Восточном Мурмане половая зрелость особей наступает чаще всего на третьем году жизни и лишь в наиболее благоприятные годы она может наступать на втором году (Кузнецов, 1953), а продолжительность жизни достигает 13 лет; лишь изредка можно встретить особей в возрасте 14 и более лет. В Белом море (южнее Горла) половая зрелость обычно наступает на втором году жизни, и в большинстве местообитаний особи живут до 10-13 лет. В Воронке и Горле Белого моря, а также на островах северной части Онежского залива длительность жизни в отдельных случаях может достигать 18-25 лет (например, в поселениях на нижней стороне камней северного берега острова Жижгина). На побережье Норвегии (Runnström, 1925) половая зрелость наступает так же, как и в Белом море, на втором году жизни, но после первого же размножения основная: Масса особей погибает, т. е. продолжительность их жизни здесь составляет всего лишь около 2 лет. Отдельные особи живут, по-видимому, до 3-4 лет. Далее к юго-западу, у берегов Бельгии, половая зрелость, как правило, наступает к концу первого же лета жизни (Lefevere, Leloup, Van Meel, 1956), т. е. в возрасте 3-4 месяцев. Интересно, что в юго-западной части Кандалакшского залива, по наблюдениям М. Н. Русановой (1959), в отдельные наиболее теплые годы половая зрелость у значительного числа особей наступает также в возрасте 3-4 месяцев. Тем не менее максимальная длительность жизни при этом не изменяется и в различных; местообитаниях колеблется от 7 до 13 лет.
В общих чертах цикл размножения В. balanoides на Мурмане был определен еще П. В. Ушаковым (1924), сообщившим, что осенью в мантийной полости появляются зародыши, а с ноября по март они находятся на стадии науплиуса, но заключены в оболочки. Выход науплиусов в планктон П. В. Ушаков наблюдал в апреле. Сходные результаты получены нами и в результате многолетних наблюдений на Восточном Мурмане. Развитие половых продуктов начинается здесь вскоре после массового выхода науплиусов и следующей затем линьки. В конце июля или в начале августа созревшие яйца оплодотворяются и начинается их дальнейшее развитие. Уже в октябре в выводковых камерах можно видеть хорошо развитых сегментированных зародышей, заключенных в прозрачную оболочку. В конце декабря или в январе зародыши уже имеют характерный науплиальный глаз и на этом их развитие приостанавливается. С наступлением весеннего потепления все личинки почти одновременно на огромном протяжении побережья выходят в планктон. Например, в 1937 г. это наблюдалось 5-7 мая, а уже 20 мая прибрежные камни и скалы были усеяны молодыми балянусами; в 1948 г. массовый выход личинок произошел в течение более длительного периода: с 26 марта до 15 апреля, а прикрепление циприсов в этом году замечено 19 мая. Сильные зимние шторма часто производят значительные разрушения в поселениях В. balanoides, "благодаря чему содержащиеся в мантийной полости зародыши попадают в водную толщу. Поэтому большое количество "куколок" и даже науплиусов можно видеть в планктоне сразу же после их появления в мантийной полости. Однако эти личинки не развиваются, и первые циприсы никогда не появляются ранее начала весны, т. е. конца апреля-начала мая. Утверждение М. М. Камшилова (1958) о том, что период естественного выхода науплиусов в планктон на Восточном Мурмане продолжается с января по май представляется малоубедительным.
В Белом море размножение этого вида более или менее изучено только на Карельском берегу (в районе губы Гридиной и в устьевой части губы Чупы) и в западной части Кандалакшского залива. Созревание половых продуктов начинается здесь в июне и во второй половине августа яйца откладываются в мантийную полость. Уже в сентябре или в самом начале октября зародыши находятся на стадии сегментации, и до наступления ледостава у них обычно успевает образоваться науплиальный глаз. В течение всей зимы эмбрионы чаще всего вмерзают в лед и дальнейшее их развитие, естественно, останавливается. Время выхода науплиусов в планктон в Белом море определяется двумя факторами: исчезновением ледяного припая и соленостью верхнего слоя воды. В результате этого нередко случается, что после исчезновения ледяного припая науплиусы задерживаются в выводковых сумках до середины июня из-за сильного опреснения поверхности воды. Обычно в различных частях Белого моря выход науплиусов наблюдается в конце мая - начале июня при температуре воды от -0.5 до +12°. В Кандалакшском заливе это чаще всего бывает при температуре воды от -0.1 до +3.0°. При сильном и длительном опреснении выход науплиусов может задержаться до середины июня, когда температура воды достигает уже 12-16°; в некоторые годы сильное и продолжительное зимне-весеннее опреснение вызывает массовую гибель не только науплиусов, но и вынашивающих их взрослых особей.
Возможно, что в разные годы время выхода науплиусов изменяется в Белом море в еще более широких пределах. Так, например, М. А. Виркетис (1926) сообщала, что появление в планктоне Белого моря науплиусов усоногих раков вообще, и В. balanoides в частности, падает на июль-август. По ее предположению личинки исчезают из планктона в августе.
По нашим наблюдениям, на Карельском берегу и в западной части Кандалакшского залива молодые балянусы появляются в конце июня или в июле. В некоторые годы метаморфоз личинок в планктоне задерживается, и молодь появляется только в сентябре или не появляется вовсе. Следствием всего этого служит неравномерность возрастного состава популяций беломорских балянусов и нерегулярное чередование "урожайных" и "неурожайных" поколений. В качестве примера можно указать, что в Кандалакшском заливе массовое появление молоди наблюдалось в 1945 и 1946 гг., в 1947 г. молоди не появилось вовсе, в 1948 г. появились лишь единичные особи, а в 1949 г. молоди опять не появлялось. В Онежском заливе в течение этих же лет молодь появлялась ежегодно, но в очень ограниченных количествах. В 1950 г. молодые балянусы в значительном количестве появились как в Онежском, так и в Кандалакшском заливах.
По материалам М. Н. Русановой, на южном берегу Кандалакшского залива в районе устья губы Чупы за период с 1951 по 1959 г. особенно "урожайными" были поколения 1951, 1955 и 1959 гг., а крайне "неурожайными" поколения 1956, 1957 и 1958 гг. Массовая гибель взрослых особей вместе в вынашиваемыми ими зародышами наблюдалась в мае 1958 г. и была обусловлена длительным опреснением поверхности воды в течение всей второй половины зимы и весной.
Сравнивая размножение балянусов в различных частях ареала (табл. 83), можно видеть, что по мере продвижения его на северо-восток период инкубации зародышей увеличивается с 4-5 у берегов Англии до 8 на Восточном Мурмане и до 9-10 месяцев в Белом море. Столь же широко изменчиво и время начала инкубации: в более низких широтах она начинается значительно позднее, чем в более высоких, хотя казалось бы, что более сильное летнее прогревание в низких широтах должно было бы вызывать и более ранее половое созревание. В пределах всего ареала наибольшее постоянство обнаруживает время выхода науплиусов, несмотря на разнообразие температурных условий. Лишь в Белом море наличие ледяного припая и зимне-весеннее опреснение поверхности воды задерживает выход науплиусов на 1.5-2 месяца по. сравнению с другими частями ареала. На Мурмане, как это уже было показано, несмотря на суровость термического режима, науплиусы могут выходить в планктон в марте-апреле, т. е. одновременно с тем, что наблюдается у берегов Англии.
| Место наблюдений | Месяцы | Средние месячные температуры воды, при которых протекает инкубация |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Ирландское море, район Мильпорта | 0 | 0 | 0- | * | * | * | * | * | * | * | * | +0 | 6.5-10 | Pyefinch, 1948; Tattersaal, 1920 |
| Южный берег Англии, район Плимута | 0 | 0 | 0- | 0- | * | * | * | * | * | * | * | +0 | 6-11 | Moor, 1934-1936; Bassindale, 1936 |
| Побережье Голландии | 0 | 0- | * | * | * | * | * | * | * | * | +0 | 0 | 5.5-11 | Delsman, 1917 |
| Южный берег Норвегии, район Бергена | 0 | 0 | 0 | 0- | + | + | * | * | * | +0 | +0 | 0 | 3.5-12 | Runnström, 1925 |
| Восточный Мурман, район губы Дальне-Зеленецкой |
0 | 0 | 0- | 0- | * | * | * | * | +0 | +0 | 0 | 0 | 0.1-8.1 | Наши наблюдения |
| Белое море, западная часть Кандалакшского залива |
0 | 0 | 0 | 0 | 0- | 0- | * | +0 | +0 | +0 | 0 | 0 | -0.4+13.7 | Наши наблюдения |
Примечание. +откладывание яиц, 0 инкубация, - выход науплиусов в планктон.
У северных берегов Англии выход науплиусов в планктон наблюдался в марте (Pyefinch, 1948) при средней месячной температуре воды 6.7°; в отдельные годы средняя температура воды в марте колеблется здесь от 4.0 до 7.6° (Barnes, 1956). V южных берегов Англии массовый выход науплиусов бывает в феврале-марте (Moor, 1934-1936; Bassindale, 1936), при температуре воды 7.8-7.9° (Orton, 1919). В Ливерпульском заливе науплиусы впервые появляются в планктоне в конце февраля и достигают максимума в середине апреля при температуре воды 6-10° (Hoek, 1909). В районе Бергена выход науплиусов наблюдался в марте при температуре воды около 4-6° (Runnström, 1925). У берегов Восточного Мурмана массовый выход науплиусов, как уже сказано, бывает в марте-апреле при температуре воды не выше 2-3°.
По сообщению супругов Бэрнс (Barnes Н. а. М. Barnes, 1958), в естественных условиях различных частей ареала (от Лофотенских островов и прибрежных вод Англии до берегов Канады) личинки В. balanoides держатся в планктоне от 21 до 48 суток и живут при температуре от 1 до 7°. В экспериментальных условиях 1 метаморфоз личинок нормально протекал в температурном диапазоне от 1.0 до 18.5°.
В последнее время показано, что южная граница распространения В. balanoides в общем совпадает с зимней (средней в январе) изотермой 8° , но во многих случаях этот вид живет и там, где температура воды не опускается ниже 10 (Barnes, 1958). Вообще следует иметь в виду, что, например, в Европе южная граница его поселений находится у берегов Португалии и Азорских островов, где температура воды ниже 10-12° практически не бывает. Поскольку выход науплиусов происходит всегда в конце зимы и с началом весеннего прогревания, то можно полагать, что в пределах всего ареала эта стадия размножения протекает в температурном диапазоне по меньшей мере от -0.5 до +13°. Совершенно особняком стоит Белое море, где науплиусы покидают мантийную полость при очень широком температурном диапазоне и в отдельные годы он может быть большим, чем в пределах всего ареала.
Столь же широко изменчивы и температурные условия прохождения других стадий размножения (табл. 84). Поэтому утверждение В. А. Свешникова (1959:830), что "в различных частях ареала этот балянус размножается при одинаково низкой температуре", представляется досадным недоразумением, возникшим в результате недостаточного знания имеющихся на этот счет материалов.
Размер тела и плодовитость размножающихся особей в различных местообитаниях и в различных частях ареала изменяется весьма существенно. К сожалению, известно на этот счет пока еще мало. У берегов Англии (Moor, 1934-1936) в нижнем отделе литорали индивидуальная плодовитость колеблется от 173 до 1372, составляя в среднем 479. В среднем отделе литорали - от 1160 до 5850, а в среднем 3352. У верхней границы поселений индивидуальная плодовитость возрастает еще более, изменяясь в пределах от 757 до 13110 эмбрионов и составляя в среднем 4611.
В различных местообитаниях на Восточном Мурмане средняя индивидуальная плодовитость колеблется от 743 до 3005, абсолютные колебания плодовитости находятся в пределах от 70 до 9237 эмбрионов. В Кандалакшском заливе Белого моря средняя индивидуальная плодовитость колеблется от 758 до 2289, т. е. довольно близка к тому, что наблюдается на Восточном Мурмане, но отдельные особи вынашивают здесь до 20000 эмбрионов. Обращает на себя внимание и то обстоятельство, что размеры тела беломорских особей значительно более мелкие, чем мурманских. В соответствии с этим относительная плодовитость (количество зародышей на 1 см3 тела) беломорских особей достигает весьма больших величин: если на Мурмане она не превышает 6551, то в Белом море самая меньшая величина ее составляет 7973, а наибольшая 15216 (табл. 84).
| Место наблюдений | n | Длина карапакса (в мм) | Плодовитость | Корелляция между длиной карапакса и плодовитостью (r±mr) |
Число зародышей на 1 см3 тела | Максимальный возраст (годы) |
||||
| M±m | σ±mσ | V±mv | M±m | σ±mσ | V±mv | |||||
| Восточный Мурман | ||||||||||
| Разные местообитания | 357 | 333±10 | 197±7 | 59.2±29 | 1603±74 | 1418±53 | 88.5±5.3 | +0.60±0.04 | 4809 | 13 |
| Открытые и полуоткрытые берега | 187 | 397±15 | 210±11 | 52.9±3.4 | 2188±114 | 1565±84 | 71.1±5.2 | +0.60±0.05 | 5114 | 13 |
| Закрытые берега | 170 | 264±12 | 155±8 | 58.7±4.2 | 959±60 | 775±42 | 80.8±6.7 | +0.31±0.07 | 3635 | 11 |
| Скалы на открытых и полуоткрытых берегах | 92 | 458±22 | 209±15 | 45.6±4.0 | 3005±205 | 1970±142 | 65.6±6.6 | +0.79±0.04 | 6551 | 7 |
| Каменистые россыпи на открытых и полуоткрытых берегах | 89 | 337±22 | 203±14 | 60.2±5.9 | 1408±95 | 896±64 | 63.6±6.4 | +0.22±0.10 | 4168 | 13 |
| Каменистые россыпи на закрытых берегах | 85 | 285±14 | 122±9 | 42.8±3.8 | 1232±103 | 950±73 | 77.1±8.8 | +0.42±0.09 | 4324 | 11 |
| Каменистые россыпи на закрытых и регулярно опресняемых берегах | 70 | 239±17 | 146±12 | 61.1±6.8 | 743±74 | 624±53 | 84.0±11.0 | +0.26±0.11 | 3106 | 8 |
| Белое море | ||||||||||
| Разные местообитания | 569 | 158±5 | 111±3 | 70.5±2.9 | 1539±50 | 1196±35 | 77.7±3.4 | +0.58±0.03 | 9742 | 13 |
| Открытые и полуоткрытые берега | 344 | 129±4 | 82±3 | 63.5±3.2 | 1971±46 | 854±32 | 43.4±1.9 | +0.69±0.03 | 15216 | 13 |
| Закрытые берега | 179 | 174±5 | 72±14 | 41.4±2.2 | 2051±87 | 1192±63 | 58.1±3.9 | +0.62±0.04 | 11793 | 11 |
| Скалы на открытых и полуоткрытых берегах | 211 | 147±5 | 78±4 | 53.1±3.2 | 1342±56 | 820±40 | 53.7±3.3 | +0.62±0.04 | 9126 | 13 |
| Каменистые россыпи на открытых и полуоткрытых берегах | 137 | 95±7 | 84±5 | 88.4±8.6 | 758±68 | 787±48 | 103.8±11.3 | +0.77±0.04 | 7979 | 12 |
| Скалы на закрытых берегах | 67 | 166±6 | 75±6 | 45.0±4.6 | 2289±171 | 1400±121 | 61.2±7.3 | +0.65±0.07 | 13734 | 7 |
| Каменистые россыпи на закрытых берегах | 112 | 179±6 | 69±5 | 38.7±2.9 | 1937±102 | 1077±75 | 56.6±4.8 | +0.58±0.06 | 10847 | 11 |
Отметим еще, что в Бассейне и заливах Белого моря скорость роста особей в первые годы их жизни в подавляющем большинстве случаев более высокая, чем на Мурмане. Например, в возрасте 2+ лет в различных местообитаниях Восточного Мурмана средний объем домика колеблется от 108 до 196 мм3, а в Белом море - от 78 до 346. Однако в последующие годы рост беломорских особей замедляется в значительно большей мере, чем мурманских и в результате они становятся мельче их. Например, в возрасте 5+ лет в различных местообитаниях Восточного Мурмана средний объем домика колеблется от 313 до 1716 мм3, а в Белом море - от 109 до 623 мм3. Наиболее медленно растут особи в Воронке и Горле, а также на северных берегах острова Жижгина.
Таким образом, биологическими особенностями беломорских В. balanoides следует считать:
- Снижение плотности поселений и биомассы.
- Неежегодное появление молоди, нередкие случаи массовой гибели личинок и даже взрослых форм.
- Более раннее по сравнению с особями Восточного Мурмана половое созревание и резко возрастающая относительная плодовитость.
- Меньшие, чем на Мурмане, размеры размножающихся особей.
- Длительность жизни беломорских и мурманских особей в большинстве случаев одинакова и значительно выше, чем в других частях ареала.
- Созревание половых продуктов, оплодотворение и начало инкубации эмбрионов чаще всего осуществляются при температурах, свойственных южной границе ареала этого вида. Инкубация протекает при весьма низких температурах, характерных для самых северных границ ареала, а выход личинок в планктон - при самых различных температурах, включающих в себя температурный диапазон всего ареала, а в некоторых случаях и превышающих его.
- Скорость роста особей в первые годы жизни более высокая, а в последующие годы более медленная, чем на Мурмане.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'