Биологические особенности беломорской Laminaria saccharina (L.) Lamour
В северном полушарии L. saccharina широко распространена в водах Атлантического и Тихого океанов. Кроме Восточного Мурмана и Белого моря она известна у берегов Новой Земли, Шпицбергена (Svendsen, 1959), в Карском и Чукотском морях. Обширные заросли она образует вдоль всего северного берега Европы до атлантического побережья Франции включительно (Зинова, 1953). В прибрежных водах Северной Америки L. saccharina встречается от 42 до 75° с. ш. в Атлантике (Taylor, 1957) и от 40° с. ш. до арктической Аляски в Тихом океане. Вдоль азиатского берега L. saccharina известна в Беринговом, Охотском и Японском морях, у Командорских островов и у Сахалина. В сибирских морях и у берегов Гренландии этот вид заменяется другим - L. solidungula J. Ас. (рис. 50), биологическими особенностями которого служат крайне замедленное развитие органов размножения и обусловленное этим медленное разрушение пластинки, в результате чего на одном растении она может сохраняться в течение 2-3 лет. Подробнее об этом будет сказано позднее, сейчас же рассмотрим некоторые вопросы биологии L. saccharina в районе наших исследований.
На Восточном Мурмане L. saccharina образует мощные заросли на всех более или менее защищенных от прибоя участках. Верхняя граница этих зарослей обычно располагается на 0.4-0.5 м выше линии отлива, а нижняя - на глубине 10-15 м. Наиболее плотные заросли образуются на глубине 3-4 м. В Воронке и Горле Белого моря характер распределения ламинарий сохраняется такой же, как на Мурмане. Южнее Горла более или менее постоянные заросли ламинарий имеются в сублиторали Кандалакшского залива на глубине до 8-10 м, а также вдоль Карельского берега и в северной части Онежского залива; особенно пышные заросли развиты в районе островов Жижгина и Анзерского, а также в некоторых губах и у островов западой части Кандалакшского залива. В южной половине Онежского залива, в Двинском заливе и вдоль всего Зимнего берега заросли ламинарий очень часто отсутствуют или сильно угнетены.
Биомасса L. saccharina подвержена очень резким сезонным колебаниям: в некоторых участках летом на уровне +0.4 м на 1 м2 субстрата может находиться до 230 растений с биомассой 67.8 кг; зимой число растений уменьшается здесь до 11, а биомасса до 1716 г. Уже на глубине 3 м летом имеется не более 20 растений с биомассой 15.7 кг; зимой же остается не менее 16 растений с биомассой 1008 г (Кузнецов, 1948б). Это свидетельствует о том, что сезонные изменения биомассы и плотности зарослей наиболее резки в верхних горизонтах, а с глубиной они в значительной мере затухают.
Указанные колебания биомассы обязаны своим происхождением двум явлениям: ежегодной смене листовой части слоевища, так называемой пластинки, и гибели многих растений с наступлением зимы. По мере увеличения глубины роль элиминации резко понижается и ведущее значение имеет лишь смена пластинки.
По нашим наблюдениям, молодые спорофиты появляются здесь в конце зимы и ранней весной. К началу августа общая длина их слоевища колеблется от 14 до 115 см, а вес от 0.6 до 140 г. Средний вес одного спорофита к концу первого лета жизни составляет 20.1 г, при общей длине 61 см.
В северной части Кольского залива роет молодой пластинки начинается в конце полярной ночи и наибольшей интенсивности достигает в феврале-марте; в это же время идет наиболее интенсивное разрушение старой части пластинки, остатки которой сохраняются еще до июля и даже до августа. Спороношение начинается здесь в июле и в конце августа или в сентябре оно заканчивается, и начинается разрушение пластинки. В такой же последовательности идет смена пластинки и на Восточном Мурмане (Киреева, Щапова, 1938; Тиховская, 1940; Кузнецов, 1948б).
На различных глубинах смена старой пластинки идет с различной скоростью. Например, на закрытых участках Восточного Мурмана (по наблюдениям в 1940 г.) вес молодой части пластинки в процентах от общего веса всей пластинки составлял:
| Глубина (в м) |
IV | V | VI | VII | VIII | IX | X |
| 0 | 54.7 | 84.1 | 100 | * | * | * | * |
| 3 | 78.9 | 80.7 | 82.6 | 100 | * | * | * |
| 12 | 61.9 | 64.8 | 69.3 | 69.6 | 71.4 | 73.1 | 73.2 |
Как видно, сначала смена старой пластинки быстрее идет на глубине около 3 м, а затем у границы отлива и на литорали. На глубине 12 м она идет столь медленно, что даже к октябрю старая часть пластинки составляет еще около четверти общего веса пластинки. Благодаря этому обстоятельству на ней успевает развиться довольно богатая эпифауна.
Особенностью беломорских ламинарий служит пышное развитие на них эпифауны, причем считается, что на Мурмане пластинка ламинарий вовсе свободна от нее (Дерюгин, 1928). В отношении беломорских растений подобное же явление наблюдалось и нами. Например, у растений, собранных в 1946 г. в западной части Кандалакшского залива, молодая часть пластинки составляла: в мае 67.2 и в июне 79.8% от общего веса всей пластинки (материал был собран на глубине 1 м). По-видимому, смена старой пластинки идет здесь с такой же интенсивностью, как и на глубине 10-12 м на Восточном Мурмане. В обоих случаях смена старой пластинки задерживается на время, достаточное для развития на ней эпифауны.
Наши наблюдения на Карельском берегу Белого моря показали, что здесь имеется в основном 2 экологические группы ламинарий: одна из них занимает более или менее открытые участки прибрежных вод, а другая - закрытые и преимущественно кутовые участки губ.
Рост пластинки и время ее смены на открытых участках прибрежных вод в течение 1947-1948 гг. хорошо видны из следующих данных:
| * | I | II | III | IV | V | VI | VII | VIII | IX | X |
| Общий вес всей пластинки (в г) | 187 | 151 | 165 | 130 | 181 | 248 | 209 | 288 | 272 | 252 |
| Вес молодой части пластинки (в % к весу всей пластинки) |
26.3 | 33.2 | 31.6 | 69.0 | 68.8 | 83.5 | 90.1 | 91.3 | 100 | 100 |
До апреля включительно общий вес пластинки постепенно уменьшается, следовательно, увеличение относительного веса ее молодой части в это время года осуществляется преимущественно за счет разрушения старой части. С мая по август увеличение относительного веса молодой части пластинки идет уже главным образом в результате ее роста.
Таким образом, в этом случае жизненный цикл пластинки весьма сходен с тем, что наблюдается на Мурмане, с той лишь разницей, что смена старой пластинки у беломорских растений вообще очень замедлена и осуществляется преимущественно в первой половине лета, тогда как на Мурмане она в основном завершается к началу весны.
В закрытых участках прибрежных вод характер жизненного цикла пластинки существенно изменяется и у значительной части растений он напоминает то, что известно для L. solidungula у берегов Гренландии (Jonson, 1904). Пластинка этого вида летом состоит из полностью сохранившихся частей, выросших в течение 2-3 предшествующих лет и остатков пластинки четвертого года жизни. Иными словами, длительность жизни пластинки в этом случае увеличивается по крайней мере до 2-3 лет, что может быть обусловлено снижением общего количества тепла, в результате чего рост идет более или менее нормально, а развитие воспроизводительных органов крайне замедляется. А так как разрушение пластинки обычно следует после спороношения, то становится понятным, почему она здесь сохраняется столь долго. Обращает на себя внимание то обстоятельство, что эта форма ламинарии распространена, по существу, исключительно там, где имеется зимний ледяной припай (рис. 50); правда, в Белом море она считается отсутствующей.
Между тем, по данным Е. С. Зиновой (1941), рост пластинки L. saccharina в Белом море заканчивается в июне-июле, а развитие спороносной части начинается в сентябре и заканчивается лишь к июню следующего года, когда завершается рост более молодой части пластинки. После выхода спор пластинка начинает разрушаться и полностью исчезает лишь в конце будущей зимы. Таким образом, в этом случае жизненный цикл пластинки беломорских ламинарий, в отличие от мурманских, заканчивается не в 6-7 месяцев, а по меньшей мере в течение 18 месяцев; полное разрушение пластинки на Мурмане завершается (на глубине до 3-5 м) за 18-20 месяцев, а в Белом море это продолжается в течение 24-28 месяцев.
Наши наблюдения в районе губы Гридиной на Белом море вполне подтвердили правильность этих указаний Е. С. Зиновой.
Все это позволяет высказать предположение о том, что беломорские L. saccharina, растущие на закрытых участках Карельского берега, представляют собой переходную форму между этим видом и L. solidungula. Что же касается такого признака, как замена ризоидов L. saccharina дисковидной подошвой у L. solidungula, то беломорские растения могут прикрепляться к субстрату как при помощи ризоидов, так и дисковидной подошвы: все зависит от характера субстрата и других внешних факторов. Интересно, что растения этого вида, подобные беломорским и сходные с L. solidungula, встречаются и в западной части Балтийского моря в районе острова Борнхольм (Rosenvinge, Lund, 1947).
Рост молодой части пластинки беломорских ламинарий идет также значительно медленнее, чем у мурманских. Например, в феврале рост молодой части пластинки на Мурмане идет со скоростью 50, а в Белом море - не более 28.5 м/г в сутки. В период летнего максимума молодая пластинка мурманских ламинарий растет со скоростью 38. а беломорских - только 17 мг/г в сутки (рис. 51).
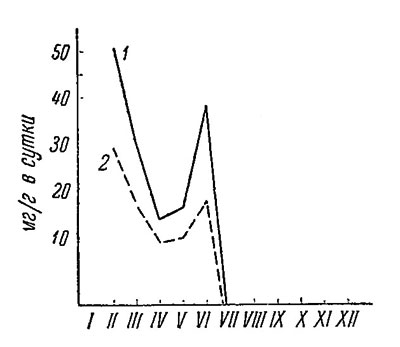
Рис. 51. Скорость роста молодой части пластинки Laminaria saccharina (мг/г в сутки) на глубине 1 м. 1 - на Восточном Мурмане (1940 г.), 2 - у Карельского берега Белого моря (1947 г.)
На Восточном Мурмане самый крупный экземпляр L. saccharina летом имел вес 2158, в Воронке Белого моря (по Мейеру, 1933) 1500, в западной части Кандалакшского залива 1309, и у Карельского берега в районе губы Гридиной 814 г*.
* (К. П. Гемп и З. Г. Паленичко (1956) сообщают, что длина спорофита ламинарий достигает "5 м и более". Из брошюры неясно, относятся ли эти цифры к беломорским растениям, или они характеризуют вид вообще. Во всяком случае, мы столь крупных растений в Белом море пока еще не встречали.)
Таким образом, биологические особенности беломорских L. saccharina состоят в сокращении ареала, уменьшении плотности зарослей, размеров наиболее крупных растений и скорости роста, а также в удлинении жизненного цикла пластинки.
В целях сравнения приводим некоторые сведения о характере жизненного цикла спорофита L. saccharina в прибрежных водах Англии, воспользовавшись для этого имеющимися литературными сведениями (Park, 1948). Основная масса зарослей образуется здесь спорофитами, появившимися весной, и лишь в закрытых участках губ к ним примешивается почти равное количество летних растений. Длительность жизни спорофита никогда не превышает 3 лет.
Рост идет в течение круглого года, но имеются ясно различимые сезонные колебания скорости роста, благодаря чему на поперечном срезе черешка можно обнаружить "годовые" кольца*. Наиболее быстрый рост протекает с января по июнь, а с июля по декабрь рост сильно замедлен. Своих максимальных размеров слоевище достигает уже во втором сезоне роста. Весьма характерной особенностью здесь является то, что, в отличие от прибрежных вод Мурмана и Белого моря, верхняя часть пластинки сбрасывается непрерывно в течение всей жизни спорофита. Нормальный возраст тканей пластинки обычно не превышает 5-7 месяцев; части пластинки в возрасте 9 месяцев не обнаружены вовсе.
* (В прибрежных водах Восточного Мурмана и Белого моря, где продолжительность жизни спорофитов, по предварительным данным, достигает 7-8 лет и более, "годовых" колец ни на черешке, ни на ризоидах обнаружено не было. Здесь о продолжительности жизни можно судить лишь путем наблюдений на контрольных площадках. Ясно различимые "годовые" кольца обнаружены пока только на поперечном срезе черешка Alaria esculenta. )
Спороношение наступает у растений в возрасте 8-12 месяцев.
Средний вес спорофитов в декабре-январе составляет 60, а в июле 310 г.
| Место наблюдений | Месяцы | Длительность созревания рецептакул (месяцы) |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Южная Англия | + | + | + | + | + | + | + | + | + | + | + | + | 9* | Parke, 1948 |
| Побережье Голландии | + | + | + | * | * | * | * | * | * | + | + | + | 6 | Van Goor, 1923 |
| Юго-запад Скандинавии | + | + | * | * | * | * | * | * | * | * | + | + | 4 | Sundene, 1953 |
| Западная часть Балтийского моря | + | * | * | + | * | + | + | + | + | + | * | + | 8 | Rosenvinge Lund, 1947 |
| Восточный Мурман | * | * | * | * | * | * | + | + | + | * | * | * | * | Киреева, Щапова, 1938 |
| Бассейн и заливы Белого моря | + | + | + | + | + | + | + | + | + | + | + | + | * | Зинова, 1941 |
* (Выход зооспор - VI-VII и X.)
Примечание. + время спороношения.
В различных частях ареала длительность спороношения колеблется от 3 до 12 месяцев; в низких широтах спороношение наблюдается преимущественно зимой, в Балтийском и Белом морях, почти в течение круглого года, а на Мурмане лишь в течение 3 месяцев (табл. 36).
Приведенные данные свидетельствуют о постепенном затухании многих биологических процессов у ламинарий по мере их продвижения от прибрежных вод Англии в Белое море. Напомним некоторые из этих данных. Длительность жизни спорофита в прибрежных водах Англии составляет 3 года, а в прибрежных водах Восточного Мурмана и Белого моря не менее 6-8 лет; жизненный цикл пластинки в прибрежных водах Англии завершается обычно в течение 5-7 месяцев, в прибрежных водах Восточного Мурмана за 18-20 и в Белом море 24-28 месяцев; в прибрежных водах Англии рост пластинки идет в течение круглого года, а на Восточном Мурмане и в Белом море рост полностью останавливается на период с августа до декабря.
В конце периода интенсивного роста (в мае-июне) средний вес одного спорофита в прибрежных водах Англии составлял 310 г, тогда как в прибрежных водах Восточного Мурмана он достигает 1050 г и более. Выше было сказано, что максимальные размеры беломорских растений немного уступают мурманским, это же наблюдается и в отношении средних размеров. Например, в западной части Кандалакшского залива средний вес взрослых растений составляет 760 г при общей длине слоевища в 182 см; интересно, что ширина пластинки в этом случае составляет 152 см. В районе губы Гридиной на Карельском берегу средний вес одного спорофита в начале лета не превышал 229 г при длине слоевища в 187 см; зимой спорофиты более крупные, и их вес составляет в среднем 477 г при длине 298 см. Таким образом, беломорские спорофиты по сравнению с мурманскими оказываются более мелкими, а по сравнению с североморскими - более крупными.
Обращает на себя внимание то обстоятельство, что, в отличие от фукоидов, размеры которых в Белом море чаще всего были крупнее, чем на Мурмане, у L. saccharina, напротив, размеры беломорских спорофитов уступают мурманским. Нельзя не заметить и того, что экологические ареалы Fucus vesiculosus и Ascophyllum nodosum в Белом море по сравнению с Мурманом резко увеличиваются, а у Laminaria saccharina сильно" сокращается.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'