Биологические особенности беломорских Ascophyllum nodosum (L.) Le Iolis
От Белого моря и Восточного Мурмана A. nodosum вдоль берегов Европы доходит до Португалии включительно. В прибрежных водах Новой Земли, Земли Франца Иосифа и Шпицбергена этот фукоид уже не встречается (Зинова, 1953). Имеется он вокруг Исландии (Jonson, 1912), у южных берегов Гренландии и вдоль атлантического берега Северной Америки от Нью-Джерси до Баффиновой Земли (рис. 47). Дрейфующие растения встречаются до Бермудских островов (Taylor, 1957).
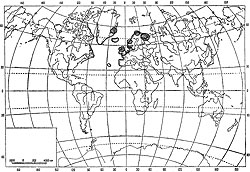
По широте своего распространения на литорали Восточного Мурмана и в Белом море этот фукоид может быть поставлен рядом с Fucus vesiculosus. Он в одинаковой мере распространен как в затишных, так и в прибойных участках, и даже там, где прибой достигает максимальной силы, можно найти отдельные низкорослые кусты этой водоросли. По-видимому, A. nodosum сравнительно легко выдерживает очень широкие колебания абиотических факторов внешней среды, но не может противостоять другим видам водорослей, становящихся часто биологически сильнее его. Лишь там, где условия внешней среды не дают возможности для жизни других видов водорослей, неизбежно появляются заросли A. nodosum, чаще всего заменяющие собой F. inflatus и F. serratus, а в верхних горизонтах литорали и F. vesiculosus. Именно поэтому A. nodosum встречается на участках, совершенно противоположных по условиям жизни в них: на скалах в верхнем отделе литорали кутовых участков губ, на скалах и камнях в затишных и сильно опресняемых участках, на скалах и камнях открытых берегов и в сублиторали Белого моря.
В Белом море (южнее Горла) этот вид распространен наравне с F. vesiculosus вдоль всех побережий, кроме Летнего и Зимнего, и образует мощные заросли как на литорали, так и в сублиторали.
В затишных участках литорали Восточного Мурмана максимальная биомасса, достигает 42, а на каменистых россыпях полузакрытых участков она поднимается до 69 кг/м2; на валунно-каменистых россыпях открытого берега биомасса может составлять до 60 кг/м2. В Воронке и Горле Белого моря максимальная биомасса колеблется на различных участках от 27.6 до 43.2, а в Кандалакшском заливе - не превышает 24.1-33.7 кг/м2. В Онежском заливе биомасса иногда может достигать 52-60, но чаще всего не превышает 30-35 кг/м2.
Время заложения молодых рецептакул в различных частях ареала может изменяться, но всегда наступает сразу после сбрасывания старых, а иногда даже предшествует ему.
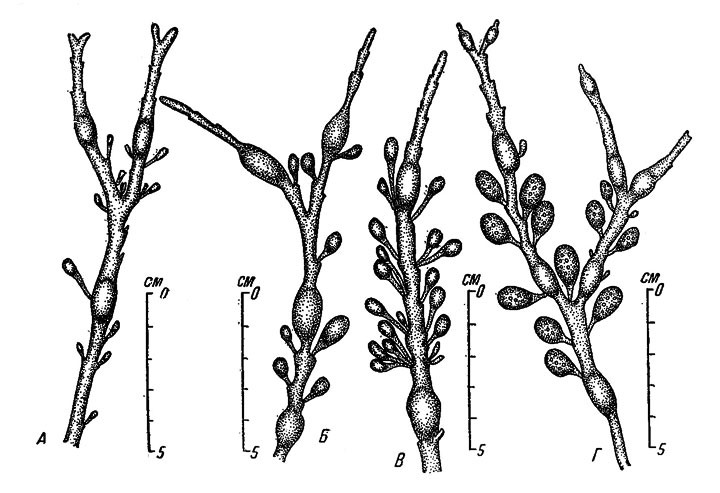
Рис. 48. Сезонные изменения внешнего вида верхней части слоевища Ascophyllum nodosum на литорали закрытых участков Восточного Мурмана. А - 6 X 1950, Б - 27 I 1951, В - 2 IV 1951, Г - 23 VI 1951
Осенью по всей длине слоевища литоральных растений Восточного Мурмана располагаются мелкие грушевидные зачатки рецептакул (рис. 48, А), которые в течение зимы постепенно увеличиваются в размерах и к началу лета достигают зрелости (рис. 48, Б, В, Г). Вскоре после этого половые продукты выходят в воду, и в течение очень короткого времени рецептакулы сбрасываются одновременно на громадных пространствах побережья. Уже в июле растения обычно полностью освобождаются от старых рецептакул и начинается развитие новых. Самые верхние из вновь развивающихся рецептакул никогда не располагаются выше последнего плавательного пузыря, образующегося в первой половине нюня (рис. 48, Г); в другое время года плавательные пузыри не образуются. Характерно дихотомическое "ветвление" слоевища - также один раз в году, в сентябре (рис. 48, А); оно предшествует остановке роста на весь зимний период. Таким образом, можно констатировать, что на Восточном Мурмане:
- Развитие новых рецептакул начинается с момента сбрасывания старых (иногда даже раньше) и продолжается в течение 11-12 месяцев. Сбрасывание отмерших рецептакул завершается в очень короткое время.
- Плавательные пузыри образуются на верхних концах слоевища один раз в году, в июне. Это дает возможность определения продолжительности жизни и скорости роста почти каждого растения.
- Дихотомическое "ветвление" слоевища происходит не на всех кончиках и также только один раз в году, в сентябре. Это служит дополнительным надежным средством определения возраста и скорости роста растений.
В Белом море в общем сохраняется такой же годовой цикл, как и на Мурмане, с той лишь разницей, что в Горле рецептакулы сбрасываются не ранее второй половины августа, а южнее его - в конце июня или в первые дни июля. Кроме этого, в южной части Онежского залива вновь образующиеся рецептакулы развиваются настолько быстро, что уже к концу августа в них имеются хорошо развитые концептакулы (рис. 49); на Мурмане это наблюдается не ранее апреля будущего года.
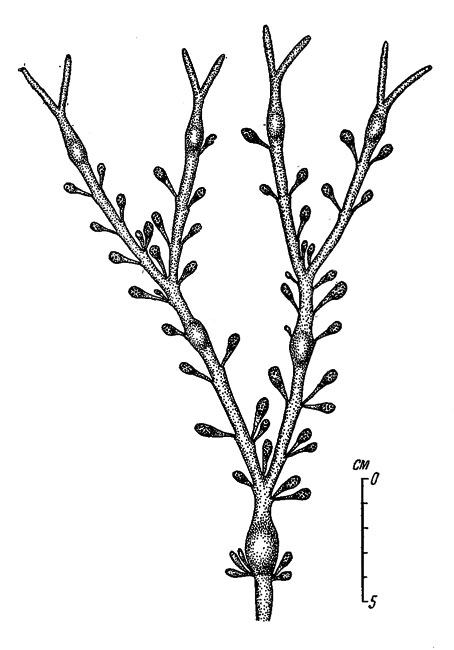
Рис. 49. Верхняя часть слоевища Ascophyllum nodosum (южная часть Онежского залива, август 1 950 г.). На молодых рецептакулах хорошо различимы скафидии (концептакулы)
Впрочем, и сбрасывание рецептакул происходит в южной части Онежского залива значительно раньше, и уже во второй половине июня растения всюду лишены их.
Органы плодоношения составляют значительную часть общего веса растения, и при сбрасывании их в водоем поступает большое количество органического вещества: На 1 кг водоросли образуется от 3 до 8.5 тыс. рецептакул весом от 110 до 530 г. В среднем вес рецептакул составляет 40-45% от общей биомассы водоросли в период зрелого плодоношения.
Сравнение некоторых биологических свойств растений с различных участков Восточного Мурмана и Белого моря (табл. 34) позволяет сделать следующие заключения:
- Максимальная длина растений на Восточном Мурмане составляет 126, а южнее Горла 153 см.
- Наибольшая продолжительность жизни растений на Восточном Мурмане равна 18+, а южнее Горла - не более 15+ лет.
- Средний годовой прирост длины слоевища на Восточном Мурмане достигает 70, а южнее Горла 91 мм*.
- Максимальная биомасса растений на Восточном Мурмане достигает 69, а южнее Горла Белого моря - не более 42.8 кг/м2.
* (Не указывая источников, Л. А. Зенкевич (1947:155) сообщает, что в Кольском заливе длина отдельных плетей A. nodosum достигает 1 м, а в Кандалакшском заливе только. 0.5 м. По нашим материалам, как это видно из табл. 28, в Кандалакшском заливе длина слоевища этой водоросли достигает 121 см, т. е. значительно больше величины, показанной Л. А. Зенкевичем для Кольского залива.)
Как видно, основными биологическими отличиями беломорских растений от мурманских служат их более крупные размеры, меньшая продолжительность жизни, более высокая скорость роста и меньшая плотность зарослей.
Жизненный цикл этой водоросли изучался и в средней части побережья Норвегии (Printz, 1926), где она является наиболее массовым представителем литоральной флоры.
Средние месячные температуры воды здесь колеблются от 3.6 в феврале до 14.5° в июле. Абсолютный температурный максимум (в июле) составляет 17.3, средняя годовая температура поверхности воды 7.9°. Соленость воды зимой более высокая, чем летом, и колеблется от 30.2 до 32.3, в июне соленость минимальная и не превышает 22.9-23.8%, в последующие месяцы она постепенно повышается и к зиме достигает указанного максимума.
| Место наблюдений | Длина наиболее крупных растений (в мм) |
Средний прирост в длину за год (в мм) |
Максимальный возраст (годы) |
Среднее количество концептакул в одной рецептакуле |
Максимальная биомасса (в кг/м2) |
| Восточный Мурман Подвергающиеся резкому опреснению кутовые участки губ |
660 | 49 | 11+ | - | 46.2 |
| Не подвергающиеся резкому опреснению кутовые участки губ | 1020 | 70 | 11+ | - | 69.0 |
| Каменистые и валунные россыпи на полуоткрытых берегах | 1260 | 50 | 18+ | - | 41.1 |
| Каменистые россыпи на открытых берегах | 400 | 33 | 8+ | 90 | 28.1 |
| Белое море. Горло Каменистые россыпи на открытых берегах |
650 | 42 | 11+ | 97 | 43.2 |
| Онежский залив Побережье островов южной части залива |
1530 | 90 | 14+ | - | 52.1 |
| Побережье островов средней части залива. При отсутствии прибоя. В верхнем отделе литорали |
580 | 60 | 10+ | - | 29.7 |
| Там же. У нижней границы отлива | 1020 | 81 | 12+ | - | 42.8 |
| Побережье островов средней части залива. При слабом прибое. В верхнем отделе литорали |
610 | 66 | 9+ | - | - |
| Там же. У нижней границы отлива | 1050 | 68 | 14+ | - | - |
| Закрытые участки Поморского берега | 1100 | 72 | 15+ | * | - |
| Открытые участки Лямицкого берега | 950 | 55 | 13+ | * | - |
| Открытые участки побережья островов северной части залива |
790 | 91 | 8+ | * | 32.5 |
| Закрытые участки побережья островов северной части залива |
440 | 61 | 6+ | * | 33.7 |
| Кандалакшский залив Закрытые участки Карельского берега |
1210 | 89 | 15+ | * | 32.5 |
| Полузакрытые участки Кандалакшского берега | 950 | 71 | 12+ | * | 28.0 |
В этих условиях плавательные пузыри, так же как на Мурмане и в Белом море, образуются только один раз в году, в марте, т. е. на три месяца раньше по сравнению с районом наших исследований. Образование плавательных пузырей непосредственно предшествует периоду интенсивного роста. Первые плавательные пузыри образуются не ранее, чем на третьем году жизни растения, поэтому возраст их определялся автором по числу плавательных пузырей плюс 2 года. Основная масса растений в Трондхьемфиорде живет 10-13 лет, но отдельные экземпляры живут до 19 лет и достигают 2 м длины. В наших водах, как уже сказано, примерно при такой же длительности жизни, растения имеют длину не более 126 на Мурмане и 153 см в Белом море.
Плодоношение в Трондхьемфиорде начинается не ранее, чем в возрасте 2 лет; по достижении половой зрелости растения размножаются еще в течение 5 лет, после чего способность к размножению утрачивается, и в возрасте 6 лет и старше рецептакул они уже никогда не имеют. Это весьма интересный факт, так как в наших водах растения не теряют способности к размножению до самого последнего года жизни. В наших водах по мере увеличения возраста растений замедляется скорость роста, но воспроизводительные способности сохраняются, в норвежских же водах скорость роста заметно увеличена, но способность к размножению с возрастом затухает. К сожалению, мы пока не располагаем достаточным материалом для подробного рассмотрения и правильной биологической оценки подобных фактов.
| Место наблюдений | Месяцы | Длительность созревания рецептакул (месяцы) |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Восточная Гренландия | * | * | * | * | * | 0 | 0 | 0 | 0 | * | * | * | 9(?) | Jonson, 1904 |
| Фарерские о-ва | * | * | * | * | * | 0 | 0 | 0 | * | * | * | + | 6 | Jonson, 1912 |
| Юго-запад Скандинавии | * | * | 0 | 0 | 0- | 0+- | +- | + | * | * | * | * | 9 | Kylin, 1907, Sundene, 1953 |
| Побережье Голландии | 0 | - | - | - | - | * | + | + | * | * | * | * | 5 | Van Goor, 1923 |
| Средняя Норвегия | * | * | 0 | 0 | 0 | 0+ | 0+- | * | * | * | * | * | 9 | Printz, 1926 |
| Восточный Мурман | * | * | * | * | * | 0+- | +- | * | * | * | * | * | 12 | Наши наблюдения |
| Северная часть Горла Белого моря | * | * | * | * | * | * | 0+ | 0+ | -+ | -+ | * | * | 12 | Наши наблюдения |
| Бассейн и заливы | * | * | * | * | * | * | 0+- | +- | * | * | * | * | 12 | Наши наблюдения |
Примечание. + заложение рецептакул, 0 зрелые рецептакулы, - сбрасывание рецептакул.
О биологии этого вида в других частях ареала имеются лишь весьма ограниченные и отрывочные сведения. Можно лишь указать, что длительность периода созревания рецептакул колеблется от 5-9 месяцев в умеренных широтах до 12 месяцев на Мурмане и в Белом море. Молодые рецептакулы на севере образуются до сбрасывания старых, а период зрелого плодоношения весьма краток - не более месяца, тогда как в умеренных широтах рецептакулы нередко образуются лишь спустя несколько месяцев после сбрасывания старых, а период зрелого плодоношения растягивается на 3-5 месяцев (табл. 35).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'