Некоторые сведения о биологических особенностях других видов беломорских водорослей
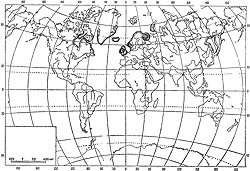
Laminaria digitata (L.) Lamour. встречается только в Атлантике и более или менее массовые заросли образуют от 30-40° с. ш. до Шпицбергена, Новой Земли и Карского моря на северо-востоке (Зинова,. 1953) и от залива Массачусетс до Гудзонова залива вдоль берегов Северной Америки (Taylor, 1957). В южных широтах заросли этого макрофита; развиваются не только в сублиторали, но и на литорали, тогда как на севере и на северо-востоке - только в сублиторали. И вообще на юге-заросли его более постоянны и непрерывны, а в высоких широтах они встречаются лишь отдельными пятнами. Восточный Мурман и Белое море служат, таким образом, одной из северо-восточных границ сравнительно небольшого ареала этого вида (рис. 52).
На Восточном Мурмане L. digitata часто встречается совместно с L. saccharina, а самостоятельные и густые заросли образуют лишь пятна вдоль более или менее открытых участков побережья. На 1 м2 дна может быть до 20 взрослых растений с биомассой до 21 кг. Вес отдельных растений достигает здесь 1.5-2 кг (Кузнецов, 1946).
Рост молодой пластинки начинается в январе и заканчивается в июле, в это же время развиваются спорангии, а в августе пластинка уже начинает разрушаться. Полное разрушение старой части пластинки обычно заканчивается не позднее мая-июня, но в некоторых случаях значительная часть старой пластинки сохраняется еще в августе и даже в сентябре (на глубинах более 3-5 м в затишных участках). Иными словами, длительность жизни пластинки, с момента начала ее роста до-полного исчезновения, составляет около 18-21 месяца.
Появление молодых спорофитов замечено в марте-апреле.
В Воронке и в северной части Горла Белого моря характер зарослей этого вида в общих чертах сохраняется таким же, как на Восточном Мурмане.
Наблюдения у Карельского берега Белого моря и на островах северной части Онежского залива показали, что здесь L. digitata встречается почти исключительно вдоль открытых берегов на глубине 1-12 м, а в губах практически отсутствует вовсе. Обычно она растет совместно с L. saccharina, Fucus serratus, а иногда и с Phyllaria dermatoidea (De la Pyl.) Le Jolis. Вообще в Бассейне и заливах Белого моря L. digitata неможет быть причислена к широко распространенным и массовым макрофитам, как на Мурмане; "чистых" зарослей этого вида в Белом море пока не обнаружено.
Самые крупные беломорские растения никогда не превышали веса 1-1.1 кг при общей длине слоевища до 90-95 см.
Рост молодой части пластинки начинается в декабре-январе и заканчивается в июне-июле. До вскрытия ледяного покрова молодая часть пластинки составляет не более 27.5-28.5% общего веса всей пластинки.
Основная часть пластинки вырастает после вскрытия льда за период с апреля-мая до июня. Тем не менее, еще в сентябре и октябре сохраняется значительная часть старой пластинки: у молодых растений старая пластинка в октябре составляет около 30% общего веса всей пластинки, а у взрослых 65-70%. Полное разрушение старой пластинки часто происходит не ранее января-марта, т. е. длительность ее жизни составляет здесь 24-27 месяцев.
У Карельского берега Белого моря спороносная часть начинает развиваться в сентябре и достигает зрелости к июню следующего года, после чего зрелое спороношение продолжается до конца жизни пластинки, т. е. по меньшей мере до конца лета, а чаще всего до весны следующего года (Зинова, 1941). В Каттегате зрелое плодоношение длится с мая до декабря (Rosenvinge, Lund, 1947). В прибрежных водах Голландии, т. е. в центре европейской части ареала этого вида, спорангии развиваются - с октября до января, но иногда они появляются еще в сентябре и сохраняются до июня (Van Goor, 1923). Здесь жизненный цикл пластинки завершается, как это видно, в более короткий срок: не более 12-18 месяцев.
Для сравнения укажем еще, что у западных берегов Скандинавии (Sundene, 1953) рост молодой пластинки начинается еще осенью, в сентябре-октябре, а к маю смена всей пластинки обычно заканчивается, в эти примерно сроки осуществляется смена пластинки и у берегов Исландии (Jonson, 1912). В Каттегате рост молодой пластинки начинается в октябре-декабре, и к весне смена старой пластинки обычно заканчивается. Однако значительная часть растений, особенно у нижней границы зарослей, меняет пластинку не ранее сентября, т. е. в этом случае длительность жизни пластинки примерно такая же, как на Мурмане, т. е. около 18-21 месяца.
Таким образом, имевшиеся в нашем распоряжении материалы свидетельствуют, что беломорские L. digitata отличаются от мурманских меньшими размерами слоевища, меньшей плотностью зарослей и значительным увеличением продолжительности жизни пластинки. Что касается общей продолжительности жизни спорофита, то в этом вопросе еще не все ясно. На Мурмане спорофиты живут, по-видимому, не менее 6-7 лет (Кузнецов, 1946), а в большинстве участков Бассейна и северной части Онежского залива - не более 4-5 лет. В юго-западной части Кандалакшского залива возраст растений, определенный по годовым кольцам на поперечных срезах черешков, достигал 10-12 лет. На побережье Норвегии (Трондхьемфиорд) растения имели возраст 10-12 и иногда 15 лет (Printz, 1926).
Alaria esculenta (L.) Grev. вдоль обоих берегов Северной Атлантики от Новой Земли и Шпицбергена опускается примерно до 40° с. ш. Должно быть отмеченным, что истинное систематическое положение многих других видов этого рода еще не вполне ясно. Возможно, что некоторые формы Alaria, считающиеся самостоятельными видами, представляют собой лишь экологические формы A. esculenta. Поэтому действительный ареал этого вида пока еще нельзя считать точно определенным.
На открытых и полуоткрытых участках литорали и верхнего отдела сублиторали Восточного Мурмана A. esculenta иногда образует довольно густые заросли, в которых другие водоросли, если и имеются, то в небольшом количестве. На закрытых участках побережья алярия часто встречается среди зарослей ламинарий ниже линии отлива. На 1 м2 дна бывает до 10 взрослых растений с биомассой до 1350 г.
Возраст каждого растения легко определяется по годовым кольцам на поперечном срезе нижней части черешка. По нашим наблюдениям, максимальный возраст растений не превышает 7+ лет, большинство же растений имеют возраст до 3 лет. К концу первого года жизни растения имеют черешок длиной 12-19 и пластинку длиной 18-80 см. Самые крупные растения (в возрасте 7+ лет) имели черешок длиной 46 и пластинку длиной 160 см.
Рост пластинки и черешка идет почти непрерывно в течение всей жизни, так же непрерывно идет и разрушение верхней части пластинки. Осенью и в первой половине зимы пребладает разрушение, и размеры растения уменьшаются, а со второй половины зимы, весной и в первую половину лета разрушение замедляется и преобладает рост, благодаря чему размеры растений увеличиваются.
Спорофилы образуются на втором году жизни растения.
Уже в Воронке и в северной части Горла Белого моря заросли A. esculenta редеют, но, в общем, здесь еще сохраняется мурманский тип; распределения этой водоросли.
В Бассейне и заливах Белого моря небольшие пятна зарослей A. esculenta встречаются лишь около мелких островов и луд в северной части Онежского залива. В Кандалакшском заливе и у Карельского берега небольшое количество растений встречается только среди зарослей ламинарий и сублиторальных фукоидов. У Поморского, Лямицкого и Летнего берегов A. esculenta отсутствует, если не считать небольшого участка в районе острова Жижгина. Беломорские растения, кроме сокращения ареала и уменьшения плотности зарослей, отличаются от мурманских и меньшими размерами: растения более 1.1 м длиной нам не встречались.
У берегов Норвегии, где возраст растений, по-видимому, не превышает 2-3 лет, пластинка полностью развита только весной и летом; осенью она почти полностью разрушается, а в январе начинается рост новой пластинки. Уже в феврале новая пластинка имеет длину 20-40 см, а в апреле-мае ее рост завершается. В это же время в спорофилах появляются сорусы.
Как видно, рост пластинки A. esculenta здесь существенно отличается от того, что наблюдается у обоих видов ламинарий: у алярии пластинка практически полностью отсутствует в течение сентября-декабря, тогда как у ламинарий молодая пластинка начинает расти еще при хорошо-сохранившейся старой. На Мурмане, наоборот, у верхней границы зарослей ламинарии осенью и в начале зимы почти полностью теряют пластинку, и новая пластинка, часто начинает расти при отсутствии старой. У алярий же подобного положения нам наблюдать не приходилось: даже в самое суровое время зимы, в полярную ночь и при непрерывных штормах у алярий всегда сохранялась значительная часть пластинки.
Ahnfeltia plicata (Huds.) относится к биполярным формам. В северном полушарии она известна из Карского, Баренцева и Белого морей; у берегов Шпицбергена, Исландии, Гренландии и Арктической Америки. Вдоль берегов Европы и Америки она опускается до 35-40° с. ш., а затем слова появляется в субантарктической области Атлантики. Имеется она и в северной части Тихого океана до Чукотского моря включительно.
На Восточном Мурмане A. plicata если и встречается, то крайне редко, во всяком случае, в описке водорослей района губы Дальне-Зеленецкой, составленном З. П. Тиховской (1948), она не упоминается. К. М. Дерюгин (1915) указывает местонахождения этого вида в южной части Кольского залива и около Малого Оленьего острова. В Белом море, напротив, A. plicata широко распространена и встречается почти повсеместно, хотя по своей общей биомассе и занимает весьма скромное место: общая биомасса ее здесь примерно в 50 раз меньше биомассы ламинарий. Тем не менее, этот вид имеет важное промысловое значение как источник получения агар-агара.
Наши наблюдения, как уже сказано, касались преимущественно лишь наиболее массовых представителей фитобентоса, и поэтому сведения по биологии анфельции весьма ограничены. Некоторые вопросы жизненного цикла и распределения этого вида в Белом море освещены в работе О. С. Зверевой (1938) и в брошюре К. П. Гемп и З. Г. Паленичко (1956).
В северной части Онежского залива небольшие пятна зарослей - A. plicata, площадью 5-10 м2, располагаются преимущественно на глубине от 1-3 до 5 м. На 1 м2 таких пятен находится от 30 до 65 растений с биомассой от 860 до 1085 г. В некоторых случаях биомасса анфельции достигает 2-5 кг/м2.
Рост слоевища этой водоросли начинается в апреле-мае и заканчивается в ноябре; зимой роста не наблюдалось. Размер годового прироста длины неповрежденного слоевища в среднем составляет всего 2 см; поврежденные растения (обрываемые штормом или срезаемые при драгировках) дают годовой прирост в среднем 3-4.6 см.
Увеличение общих размеров и веса слоевища неповрежденных растений происходит путем ежегодного правильного дихотомического ветвления, наблюдаемого в конце сезона роста. Это дает возможность правильного определения возраста каждого растения, максимальной продолжительности жизни (по числу дихотомических ветвлений - каждое ветвление соответствует одному году жизни растения) и скорости роста. Кроме этого, на поперечных срезах нижней части растения бывают хорошо видны годовые кольца, по числу которых также можно определить возраст растения, на что впервые было указано еще Джонсоном в конце прошлого столетия (Jonson, 1890-1891). В настоящее время считается, что продолжительность жизни беломорских растений анфельции составляет 6-8 лет. Возможно, что максимальная продолжительность жизни растений несколько большая, так как наиболее крупные из них имеют слоевище длиной до 40 см. В Трондхьемфиорде (средняя часть побережья Норвегии), где этот вид живет от нижней части литорали до глубины 4-5 м, продолжительность жизни растений также составляет 6-8 лет (Printz, 1926).
Размножение анфельции изучено еще недостаточно. Известны только органы бесполого размножения - нематеции, формирующие моноспоры. У беломорских растений нематеции образуются осенью, сохраняются в течение всей зимы, а период спороношения в северной части Онежского залива продолжается с апреля до начала июля.
Образование и развитие нематеции не имеет какого-либо существенного значения для изменения биомассы размножающихся растений.
Приведенные здесь сведения позволяют предполагать, что годовая продукция беломорской анфельции составляет приблизительно 1/8-1/6 часть (12-17%) ее общей биомассы.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'