Биологические особенности беломорских Fucus inflates M. Vahl.
Этот весьма вариабильный вид довольно широко распространен в северном полушарии. В Атлантике он хорошо известен у берегов Англии, Франции, Швеции и Норвегии. Далее на северо-восток он доходит до Новой Земли и Карского моря включительно*. Вдоль азиатских берегов Тихого океана F. inflatus имеется в Беринговом, Охотском и Японском морях, у Камчатки, Сахалина, Командорских и Курильских островов.
* (У берегов Шпицбергена имеется большинство форм этого вида, в том числе и та, что относится вами к типичной форме, объединенных П. Сведсеном (Svendsen, 1957) в один вид - F. distichus.)
Восточный Мурман и Белое море по существу являются северо-восточной окраиной географического ареала F. inflatus и его многочисленных форм (рис. 34).
Говоря о географическом распространении этого вида, следует иметь в виду, что многими европейскими авторами он нередко объединяется с F. distichus, а американскими - нередко бывает наделен иными названиями, например, F. edentatus De la Pyl. (Powell a. Lewis, 1952). В соответствии с этим точная картина географического распространения F. inflatus пока еще не может быть установлена.
Здесь дается очерк биологии F. inflatus и несомненно близкого к нему вида или даже только формы F. distichus. Это делается в одном разделе потому, что на практике определение многочисленных "видов" и форм, объединяющихся в одну группу общностью признаков с этим широко-распространенным и весьма полиморфным видом, часто делается совершенно невозможным. Лишь детальное изучение биологии и деталей жизненных циклов в их изменчивости под влиянием условий внешней среды у различных форм этой группы даст возможность определить их действительное систематическое положение, а также экологические и географические ареалы.
Заметим еще, что недавно значительная часть "видов" и форм этой группы была объединена в один вид - F. distichus, имеющий четыре подвида: distichus, anceps, edentatus, evanescens (Powell, 1957). Все эти формы известны как на Восточном Мурмане, так и в Белом море.
В нашем очерке F. distichus нередко называется "прибойной формой" F. inflatus, так как долгое время нам не удавалось с полной уверенностью провести границы между этими двумя видами, особенно в условиях Восточного Мурмана. В Белом море F. distichus, напротив, часто экологически и морфологически хорошо обособлен от F. inflatus и поэтому здесь он называется своим настоящим именем.
Таким образом, здесь будут рассмотрены некоторые материалы по биологии формы с широкопластинчатым талломом, снабженным более или менее обширными воздушными полостями (временными, а иногда и постоянными), для которой сохраняется название F. inflatus M. Vahl. (f. latifrons) (Зинова, 1953), и близких к ней форм, объединенных X. Т. Пауэлом (Powell, 1957) в один вид под названием. F. distichus.
В отличие от F. vesiculosus представители этого вида поселяются в более низких горизонтах литорали и ниже линии отлива нередко оказываются не только в Белом море, но и на Мурмане.
Те средние и нижние горизонты литорали, которые на Мурмане заняты сплошным поясом зарослей F. inflatus, в Бассейне и заливах Белого моря (южнее Горла) заняты зарослями F. vesiculosus или Ascophyllum nodosum. Сплошные заросли - F. inflatus, подобные мурманским, обычно не идут южнее северной половины Горла. В Бассейне и заливах Белого моря лишь отдельные пятна зарослей F. inflatus можно встретить вдоль Терского берега, в Кандалакшском заливе и на островах северной части Онежского залива.
Биомасса этого фукоида на литорали Восточного Мурмана, но Т. П. Гурьевой (1948), колеблется от 714 до 1985 г/м2 субстрата. Сходные цифры приводит и Т. А. Матвеева (19486), по данным которой средняя биомасса составляет здесь 1622 г/м2 субстрата. По нашим данным, максимальная биомасса на редких каменистых россыпях закрытых участков литорали летом колеблется от 13200, до 17600 г/м2 субстрата. В Воронке и Горле Белого моря биомасса увеличивается до 24 кг/м2, а южнее Горла нигде не превышает 6.5-9.1 кг/м2.
Наглядное представление о годовом ходе жизненного цикла взрослых растений можно получить путем наблюдений за сезонными изменениями верхней части слоевища. Например, на закрытых участках литорали Восточного Мурмана верхняя часть осенних растений обычно полностью лишена каких-либо дополнительных образований или их остатков (рис. 35, А). Однако вскоре на кончиках слоевища появляются слабые утолщения (рис. 35, Б, В) - будущие рецептакулы. Эти утолщения довольно быстро увеличиваются в размерах, и в марте в них уже появляются концептакулы; в конце марта рецептакулы выглядят уже вполне зрелыми (рис. 35, Г, Д), а в конце мая - в июне половые продукты выходят в водную толщу. Разрушение рецептакул и несших их частей слоевища протекает довольно быстро, и к осени они обычно исчезают без следа.
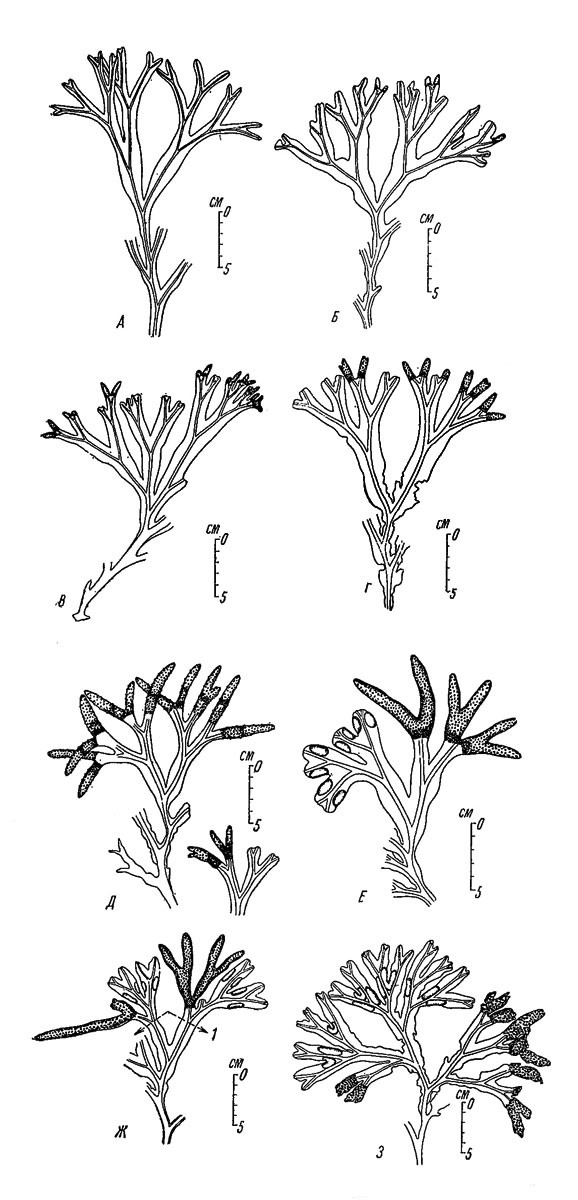
Рис. 35. Сезонные изменения внешнего вида верхней части растений Fucus inflatus на закрытых участках литорали Восточного Мурмана (губа Дальне-Зеленецкая, бухта Оскара). А - 22 X 1949, Б - 15 XII 1950, В - 9 I 1950, Г - 15 III 1950, Д - 1 IV 1950, Е - 30 IV 1950, Ж - 30 V 1950: 1 - верхние границы поселения Spiroibis bvreatis поколения 1949 г, 3 - 5 VIII 1950
Вегетативные кончики слоевища в течение всей зимы не растут, и их верхний край располагается примерно на высоте средней части рецептакул. Во второй половине апреля на вегетативных кончиках появляются плавательные пузыри (рис. 35, Е, Ж, З), которые в отличие от подобных образований у F. vesiculosus в большинство своем не имеют определенной формы и представляют собой лишь временные приспособления на период пониженной плотности воды и полностью исчезают к началу зимы. Изменения формы и размеров плавательных пузырей в течение лета можно хорошо видеть также при наблюдении в садках за одними и теми же растениями (рис. 36). После образования плавательных пузырей рост вегетативных частей слоевища идет сравнительно быстро и заканчивается в сентябре.
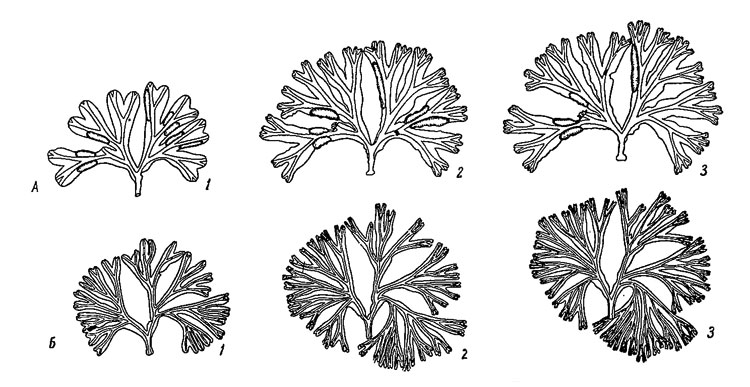
Рис. 36. Изменения внешнего вида и рост растений Fucus inflatus при содержании в садках на закрытых участках литорали. А - затишная (типичная) форма: 1 - 15 VI, 1950 10.07 г, 2 - 22 VIII 1950 15.60 г, 3 - 21 IX 1950 17.85 г. Б - прибойная (типа distichus) форма: 1 - 15 VI 1950 5.92 г, 2 - 22 VIII 1950 8.70 г, 3 - 21 IX 1950 10.25 г.
На литорали открытых участков Восточного Мурмана годовой ход. жизненного цикла взрослых растений совершается в той же последовательности, как в только что рассмотренном случае: разница состоит лишь в том, что плавательные пузыри здесь уже не образуются.
Итак, в течение года основные биологические процессы у F. inflatus распределяются следующим образом:
- Молодые рецептакулы появляются в ноябре-декабре, в марте-в них образуются концептакулы, а в мае-июне половые продукты выходят в воду. Разрушение рецептакул и несших Их, частей слоевища завершается к осени.
- Рост вегетативных частей слоевища начинается в апреле и продолжается до сентября.
- На закрытых участках литорали на вегетативных кончиках слоевища в апреле образуются крупные плавательные пузыри, исчезающие осенью.
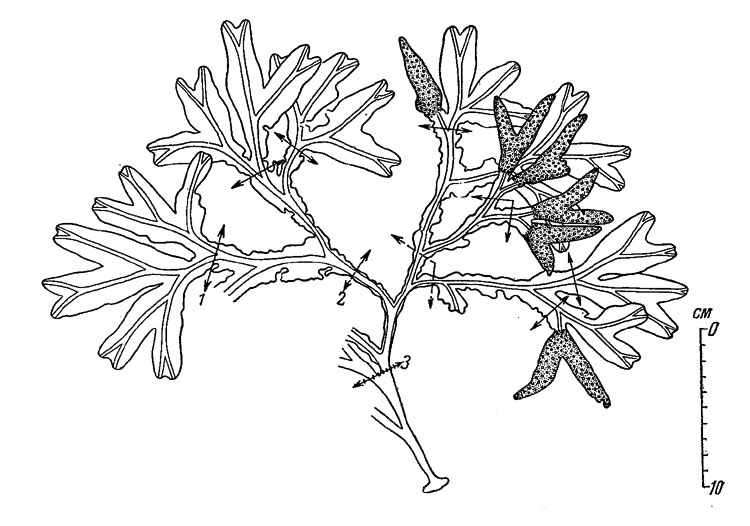
Рис. 37. Взрослое растение Fucus inflalus (Онежский залив, восточный берег о. Жижгина, 29 VI 1950). Стрелками показаны верхние границы поселений Spirorbis borealis поколения 1949 (1), 1948 (2) и 1947 гг. (3)
У взрослых растений большинство кончиков слоевища занято рецептакулами и ежегодно разрушается вместе с ними. Размер годовогоприроста длины слоевища и возраст растений у представителей этого вида можно определить тем же способом, как это делалось у прибойной формы F. vesiculosus, т. е. по числу дихотомических ветвлений слоевища от основания растения до точки ответвления его генеративных частей. Для контроля за правильностью подобных определений удобно пользоваться положением верхней границы поселений Spirorbus borealis, который поселяется на слоевище этого фукоида не только в Белом море, но и на Мурмане. Следует лишь заметить, что в Белом море прикрепление личинок этого червя происходит в конце июня или в июле, а на Мурмане - обычно в первые дни августа. По величине годового прироста, равного расстоянию между верхними границами поселений двух смежных поколений, не представляет труда приблизительно определить возраст растения (рис. 37).
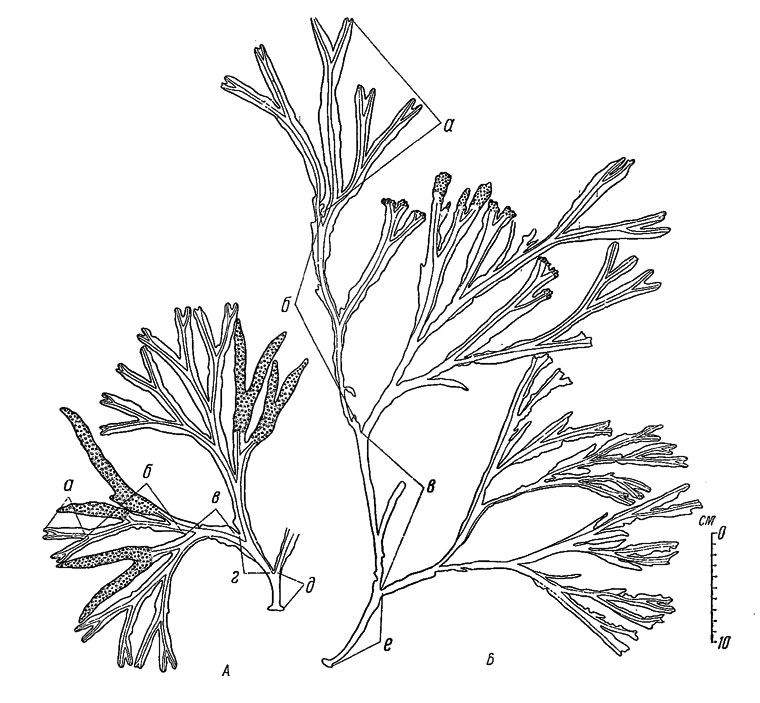
Рис. 38. Различия в скорости роста и размерах растений Fucus inflates в разных горизонтах литорали (Восточный Мурман, губа Дальне-Зеленецкая, 9 VIII 1950). А - у верхней границы зарослей, Б - у нижней границы зарослей. Годовые приросты длины слоевища: а - VI 1949-VIII 1950, б - VI 1948-VI 1949, в - VI 1947-VI 1948, г - VI 1946-VI 1947, д - после первой зимовки до VI 1946, е - после первой эимовки до VI 1947
Обращает на себя внимание, что с повышением горизонта литорали скорость роста и размеры растений уменьшаются, т. е. происходят такие же изменения, как и у F. vesiculosus. Так, например, на валунно-каменистых россыпях открытых участков литорали Восточного Мурмана у верхней границы зарослей годовой прирост длины слоевища в среднем составляет 58, а у нижней границы зарослей 180-190 мм (рис. 38). Резко замедляется рост и уменьшаются размеры растений в случаях прикрепления их к скалам (рис. 39). Например, в одном и том же пункте на литорали открытых берегов Восточного Мурмана средний годовой прирост длины слоевища у растений, прикрепленных к скале, составлял 19, а у прикрепленных к камням и валунам 105 мм. Соответствующим образом изменялись и общие размеры растений. Такие различия обусловлены, по-видимому, недостаточным питанием растений, живущих на скалах (Кузнецов, 1956).
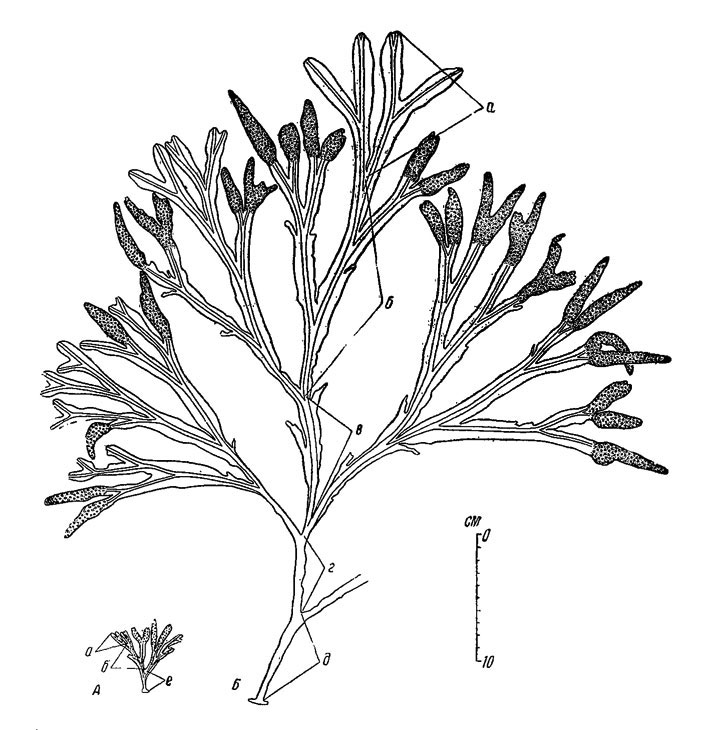
Рис. 39. Значение характера субстрата для скорости роста и размеров слоевища растений Fucus inflatus (Восточный Мурман, район губы Дальне-Зеленецкой, VII 1950). А - растение взято на скале, Б - на валунно-каменистых россыпях. Годовые приросты длины слоевища: а - VI 1949-VII 1950, б-д - см обозначения на рис. 38, е - после первой зимовки до VI 1948
Изменения размеров годового прироста дЛины слоевища, продолжительности жизни растений и некоторых других биологических свойств в популяциях Восточного Мурмана и Белого моря (табл. 30) позволяют сделать следующие заключения:
- Наибольшей продолжительностью жизни обладают растения в Воронке Белого моря (8+). Южнее Горла Белого моря растения, по-видимому, не живут более 3+, тогда как на Мурмане встречаются растения в возрасте 5+.
- Ширина "ветвей" слоевища в Баренцевом море (20 мм) меньше, чем в Белом море (24 мм). Особенно широки "ветви" слоевища в Воронке Белого моря (до 31 мм).
- Годовой прирост длины слоевища в Баренцевом море (115 мм) меньше, чем в Воронке и Горле Белого моря (до 328 мм). В Бассейне Белого моря величина годового прироста (до 124 мм) практически одинакова с тем, что имеется на Мурмане, но в Онежском заливе скорость роста заметно снижена.
- Среднее количество концептакул в одной рецептакуле наиболее высокое в Воронке Белого моря и наиболее низкое в Бассейне и в Онежском заливе.
| Место наблюдений | Средний прирост в длину за год (в мм) |
Ширина пластинки (в мм) |
Максимальный возраст (годы) |
Среднее количество концептакул в одной рецептакуле (двойной/одинарной) |
| Восточный Мурман Подвергающиеся резкому опреснению кутовые участки губ |
76 | 13 | 5+ | 608/308 |
| Не подвергающиеся опреснению кутовые участки губ | 91 | 20 | 4+ | 1088/564 |
| Каменистые россыпи на полуоткрытых берегах | 105 | 12 | 3+ | - |
| Отвесные и ступенчатые скалы на открытых берегах | 33 | 7 | 4+ | - |
| Каменистые россыпи на открытых берегах | 115 | 16 | 3+ | 468/625 |
| Опресняемые участки на каменистых россыпях открытых берегов |
36 | 5 | 2+ | 378/254 |
| Воронка Белого моря Кутовые участки губ |
75 | 18 | 3+ | 324/205 |
| Каменистые россыпи на полуоткрытых берегах | 38-125 | 15-20 | 8+ | 1109/559 |
| Валунные и каменистые россыпи на открытых берегах | 106/328 | 18-31 | 4+ | 3008/1541 |
| Скалы на открытых берегах | 19 | 3 | 2+ | 615/283 |
| Горло Белого моря Каменистые россыпи на полуоткрытых берегах |
93 | 17 | 3+ | 534/709 |
| Скалы и валунные россыпи на открытых берегах | 26 | 9 | 4+ | 1345/518 |
| Бассейн Белого моря Открытые участки южного берега |
124 | 12 | 2+ | 765/402 |
| Открытые участки северного берега | 119 | 11 | 3+ | 437/207 |
| Онежский залив Побережье островов средней части залива. При слабом прибое |
78 | 14 | 3+ | 603/356 |
| Побережье островов средней части залива. При сильном прибое |
107 | 15 | 3+ | 536/184 |
| Побережье островов северной части залива. При слабом прибое |
61 | 16 | 3+ | 644/332 |
| Побережье островов северной части залива. При сильном прибое |
74 | 24 | 3+ | - |
Таким образом, беломорские растения этого вида отличаются от баренцевоморских несколько сокращенной длительностью жизни и уменьшенной скоростью роста вегетативных частей слоевища. Характерно также значительное уменьшение удельного веса его зарослей в общей массе растений прибрежной зоны.
Относительный вес рецептакул в Белом море и на Восточном Мурмане приблизительно одинаков и составляет около половины общего веса растения в пору зрелого плодоношения.
Сведения о биологии этого вида в других частях ареала весьма скудны, и по имеющимся литературным данным можно лишь установить, что время заложения рецептакул довольно постоянно и приходится на октябрь-ноябрь, а продолжительность периода их созревания колеблется от 5 до 7 месяцев (табл. 31).
| Место наблюдений | Месяцы | Длительность созревания рецептакул (месяцы) |
Автор | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Восточная Гренландия | * | * | * | - | * | 0 | 0 | 0 | * | * | * | * | 7(?) | Jonson, 1904 |
| Юго-западная Исландия | * | * | 0- | 0- | 0- | 0- | 0- | 0- | - | + | + | * | 5 | Jonson, 1912 |
| Юго-западная Скандинавия | * | * | 0 | 0 | - | - | * | * | * | + | + | * | 5 | Sundene, 1953 |
| Восточный Мурман | * | * | * | 0 | 0 | 0 | - | - | - | * | + | + | 5 | Наши наблюдения |
| Бассейн и заливы Белого моря | * | * | * | * | * | 0- | 0- | - | - | * | + | + | 7 | Наши наблюдения |
Примечание. + заложение рецептакул, 0 зрелые рецептакулы, - сбрасывание рецептакул.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'