Биологические особенности беломорских Fucus vesiculosus
Этот вид служит одним из ведущих представителей флоры Восточного Мурмана и Белого моря и широко распространен по обоим берегам Атлантики (рис. 24). От Новой Земли и Гренландии на севере он опускается вдоль берегов Америки до Северной Каролины (Taylor, 1957), т. е. примерно от 80 до 35° с. ш. и вдоль берегов Европы и Африки до Алжира и Марокко (Зинова, 1953). Еще не так давно (Van Goor, 1923) считалось, что этот вид имеется и у берегов Южной Америки до Бразилии включительно. Хотя это указание и признано сейчас ошибочным и южно-американские фукоиды выделены в самостоятельные группы, оно, тем не менее, позволяет предполагать, что на литорали Южной Америки встречаются формы экологически и морфологически близкие к нашему F. vesiculosus.
Восточный Мурман и Белое море, где проведены наши наблюдения, расположены в непосредственной близости к северо-восточной границе ареала этого вида.
Общий характер жизненного цикла макрофитов и распределение во времени некоторых свойственных им биологических процессов можно проследить путем зарисовки контуров растений, наиболее типичных для данного местообитания и взятых в разное время года. Такого рода работа была проведена в течение 1948-1953 гг. на побережьях Баренцева и Белого морей, и частичным результатом ее служат приводимые ниже сведения о различиях в скорости роста, размерах слоевища, продолжительности жизни и времени размножения у некоторых фукоидов Восточного Мурмана и Белого моря.

Рис. 25. Сезонные изменения внешнего вида верхней части растений Fucus vesiculosus на литорали закрытых участков Восточного Мурмана. А - 22 X 1949; В - 12 XII 1949; В - 15 III 1950; Г - 1 IV 1950; Д - 30 IV 1950; Е - 30 V 1950; Ж - 20 IX 1950
На закрытых участках Литорали Восточного Мурмана на осенних растениях F. vesiculosus имеются более или менее сохранившиеся, но уже пожелтевшие и разрушающиеся рецептакулы, а на уровне их верхнего края на вегетативной части слоевища обычно расположен ряд плавательных пузырей (рис. 25, А). В последующие затем месяцы происходит окончательное разрушение старых рецептакул, а в декабре на кончиках слоевища уже различаются слабые вздутия - будущие рецептакулы. В марте эти вздутия уже имеют сходство с мелкими рецептакулами, а вскоре у них появляются и концептакулы (рис. 25, Б, В, Г). Вегетативные кончики слоевища, т. е. лишенные вновь образующихся рецептакул, начинают свой рост в наиболее холодное время года, в марте или в апреле (иногда в мае), когда на них образуется ряд плавательных пузырей (рис. 25, Д, Е). С этого времени рост вегетативных частей слоевища идет очень быстро, а рецептакулы к концу мая достигают зрелости, в течение ближайшего времени половые продукты выходят в воду и постепенно растения приобретают тот же вид, который они имели осенью предшествующего года, с той лишь разницей, что за истекший год на них прибавился один ряд плавательных пузырей (рис. 25, Ж). Иногда на отдельных растениях в течение лета могут образовываться дополнительные единичные, или даже группами, плавательные пузыри, которые всегда по своему количеству, положению и размерам хорошо отличимы от образовавшихся весной (рис. 26). Образование этих дополнительных плавательных пузырей связано с температурой и соленостью: чем сильнее летнее прогревание и чем сильнее весеннее или летнее опреснение, тем чаще и тем в большем количестве образуются эти пузыри. На Мурмане они встречаются крайне редко, только в кутовых участках губ, в Белом море - часто; обычны они и по берегам западной Европы.
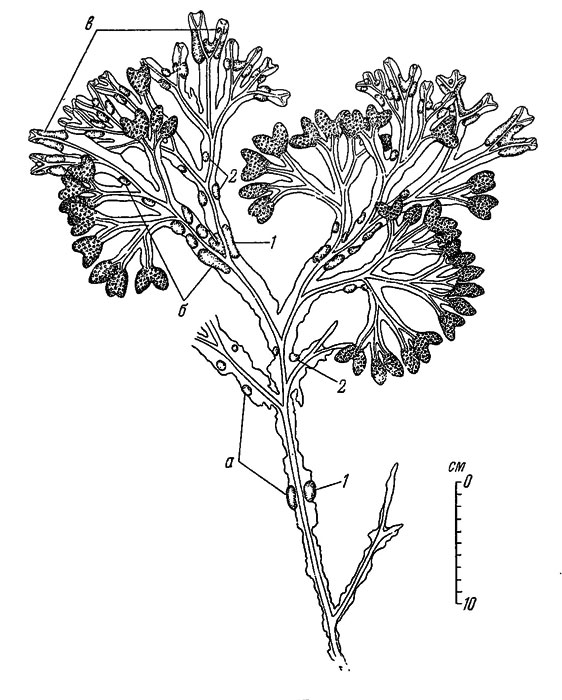
Рис. 26. Верхняя часть растения Fucus vesiculosus (глубина 1 м в Кандалакшском заливе). Группы плавательных пузырей (а, б, в): плавательные пузыри, А - около образовавшиеся весной (1) и в разное время лета (2)
Таким образом, на закрытых участках литорали Восточного Мурмана:
- Заметное простым глазом развитие рецептакул начинается в декабре, а концептакулы образуются в марте.
- Плавательные пузыри образуются, как правило, один раз в году - в апреле или в мае. Это дает возможность определять возраст растений и размер их ежегодного прироста.
- Части слоевища, непосредственно несущие рецептакулы, после окончания периода размножения полностью разрушаются и в течение осени и зимы поступают в общий круговорот веществ.
- Рост вегетативных частей слоевища начинается в марте, но до образования плавательных пузырей идет очень медленно. Основная масса слоевища создается после образования плавательных пузырей, за период с апреля-мая до сентября.
На литорали открытых берегов плавательные пузыри на слоевище F. vesiculosus обычно не образуются, и это делает затруднительным определение возраста и скорости роста растений. Кроме этого, существенной биологической особенностью растений, живущих на открытом берегу в условиях более или менее постоянного смачивания прибоем, а, следовательно и смягчения колебаний (сезонных, суточных и погодных) факторов внешней среды, служит довольно ясно выраженное нарушение в ходе годового цикла взрослых растений. Это нарушение состоит в том, что рост слоевища и созревание рецептакул могут протекать в любое время года. Однако в основной своей массе растения и в этих условиях растут и плодоносят в общем в те же сроки, что и на закрытых участках литорали. Здесь в полной мере сохраняется правило, что все генеративные части слоевища после окончания периода размножения разрушаются, и это дает возможность определять продолжительность жизни и ежегодный прирост по количеству дихотомических ветвлений, расположенных ниже точки ответвления генеративных частей слоевища. Bce, что находится выше этой точки, считается выросшим в течение года, предшествующего наблюдению.

Рис. 27. Различные формы растений Fucus vesiculosus, лишенные плавательных пузырей (литораль, Онежский залив). А - около устья реки Лямица, 28 VIII 1950; Б - юго-западный берег о. Конд-остров, 7 VII 1950
В губах Воронки, на значительном протяжении Терского берега, в Кандалакшском и Онежском заливах F. vesiculosus почти всегда снабжен нормально развитыми плавательными пузырями; растения, лишенные плавательных пузырей, встречаются здесь главным образом на скалах и камнях открытых берегов Воронки и мелких островов Онежского залива (рис. 27).
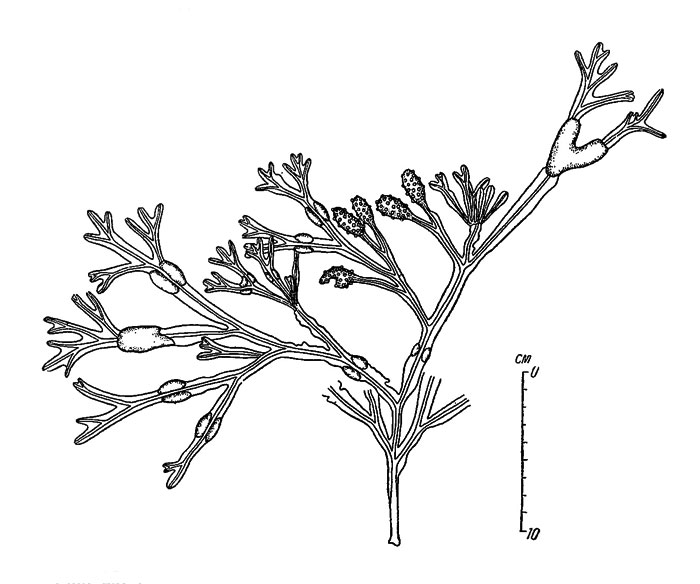
Рис. 28. Растение Fucus vesiculosus (литораль Лямицкого берега Белого моря). Показаны сильно вздутые, неправильной формы плавательные пузыри
На литорали южной части Поморского, вдоль всего Лямицкого и Летнего берегов плавательные пузыри обычно отличаются крупными размерами и нарушением их обычной формы (рис. 28). Здесь, как уже сказано, часто в течение лета образуется не один, а несколько рядов плавательных пузырей (рис. 24).

Рис. 29. Изменения внешнего вида и рост различных форм Fucus vesiculosus при содержании их в садке на закрытом участке литорали Восточного Мурмана. А - прибойная форма: 1 - 21 XI 1950 2.25 г, 2 - 23 I 1951 2.15 г, 3 - 27 II 1951 2.10 г, 4 - 20 III 1951 2.10 г, 5 - 24 IV 1951 2.75 г, 6 - 29 V 1951 3.60 г, 7 - 30 VI 1951 4.95 г, образовались плавательные пузыри, отсутствующие в естественных условиях. Б - верхняя часть растения типичной формы: 1 - 21 XI 1950 7.85 г, 2 - 14 XII 1950 8.15 г, 3 - 23 1 1951 7.70, 4 -27 II 1951 8.10 г, 5 - 27 III 1951 7.70 г, 6 - 24 IV 1951 9.47, 7 - 29 V 1951 13.10 г. В - молодые растения типичной формы: 1 - 15 VI 1950 5.75 г, 2 - 22 VIII 1950 8.60 г, 3 - 21 IX 1950 10.45 г.
Результаты наблюдений над сезонными изменениями состояния фукусов, полученные путем изучения в разное время года разных растений, полностью подтверждаются и результатами наблюдений в садках за одними и теми же растениями (рис. 29). Интересно, что при содержании в садке, расположенном в затишном участке, у прибойной формы F. vesiculosus образуются плавательные пузыри в первый же год жизни в этих необычных для нее условиях.
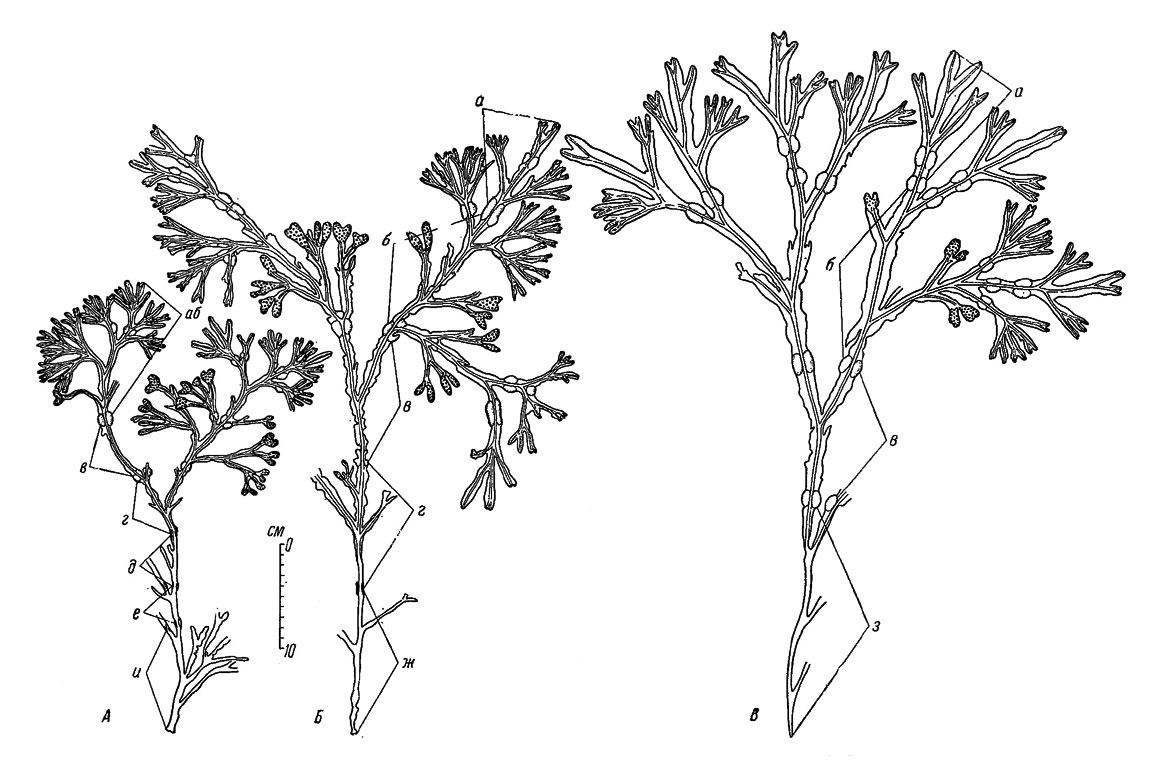
Рис. 30. Различия в размерах, скорости роста и продолжительности жизни растений Fucus vesiculosus в различных горизонтах литорали и сублиторали на Поморском берегу Белого моря (губа Колежма, Омостров, 7 IX 1950). А - верхний отдел литорали, Б - нижний отдел литорали, В - сублитораль, глубина 1 м. Годовые приросты длины слоевища: а - V-IX 1950, б - V 1949-V 1950, в - V 1948-V 1949, г - V 1947-V 1948, д - V 1946-V 1947, е - V 1945-V 1946, ж - VII 1946-V 1947, з - VII 1947-V 1948, и - VII 1944-V 1945
Обращает на себя внимание тот факт, что растения верхнего отдела литорали мельче растений ее нижнего отдела и особенно сублиторали (рис. 30). Эти различия достигают значительных величин как в Белом море, так и на Мурмане. Например, в южной части Поморского берега средний годовой прирост длины слоевища у верхней границы зарослей 62, у границы отлива 166 и в сублиторали на глубине 1 м 183 мм.
Для сравнения приводим сведения о годовых приростах слоевища F. vesiculosus в различных горизонтах литорали Восточного Мурмана:
| Характеристика участка | Уровень над 0 глубины (в см) | Годовой прирост (в мм) |
| Отдельные камни на илисто-песчанистом пляже закрытых участков. Незначительное опреснение весной. Зимой слабый ледяной припай. |
280 | 69 |
| 210 | 96 | |
| 176 | 152 | |
| Отвесные и ступенчатые скалы открытых берегов. Сильный прибой. Источников опреснения нет. |
350 | 18 |
| 280 | 29 | |
| 240 | 35 |
Самые мелкие растения, имеющие очень замедленный рост и вообще угнетенное состояние, встречаются в различного рода скалистых ваннах в верхнем отделе литорали (рис. 31). Здесь сказываются, очевидно, частые и весьма резкие колебания многих факторов внешней среды, а также ограниченное поступление минеральных питательных веществ.
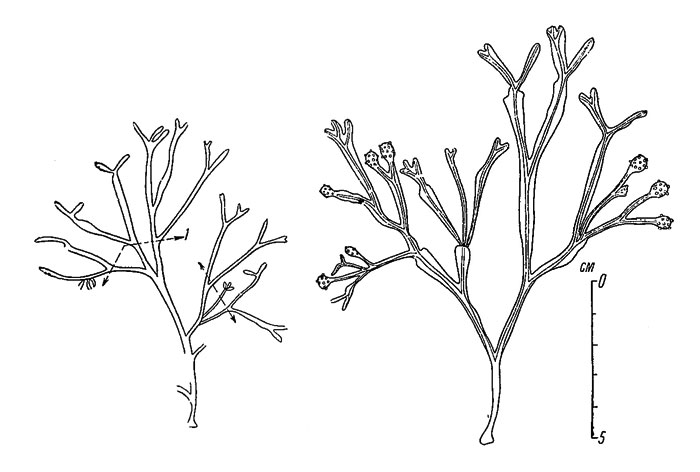
Рис. 31. Наиболее угнетенные растения Fucus vesiculosus из скалистых ванн в верхнем отделе литорали (Онежский залив, западный берег Соловецкого острова, 17 VII 1950). 1 - верхние границы поселений Spirorbis borealis поколения 1949 г.
В некоторых случаях дополнительным средством определения величины годового прироста слоевища, особенно важным там, где плавательные пузыри не образуются, у беломорских F. vesiculosus служат поселения на них многощетинкового кольчеца Spirorbis borealis. Прикрепление к поверхности слоевища личинок этого червя в Белом море происходит в конце июня или в начале июля, причем вновь поселившиеся особи занимают всю верхнюю часть слоевища. В соответствии с этим вся свободная от них верхняя часть слоевища, взятого после прикрепления личинок червя, является выросшей с указанного времени. Кроме того, на слоевище в таких случаях всегда хорошо видна и верхняя граница поселений червя поколения предшествующего года, а расстояние между верхними границами поселений двух поколений и составляет размер годового прироста водоросли. Пример подобных растений Fucus vesiculosus показан на рис. 32.
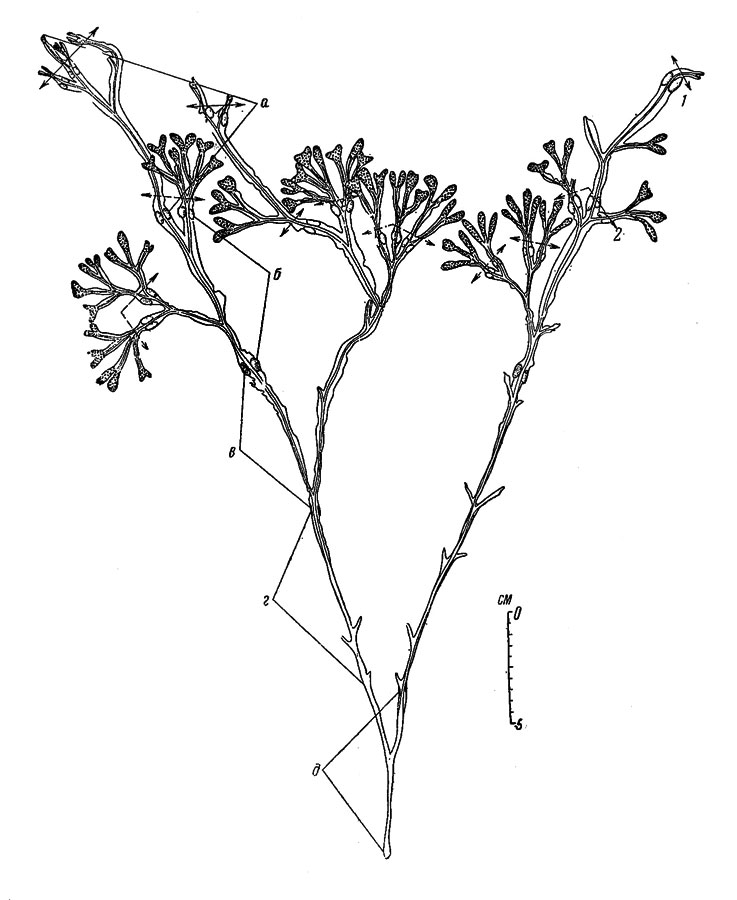
Рис. 32. Растение Fucus vesiculosus (сублитораль у западного берега о. Соловецкого, 17 VII; 1950). Верхние границы поселений Spirorbis borealis поколения 1950 (1) и 1949 (2). Годовые приросты длины слоевища: а - V 1949-VII 1950., б - VII 1945-V 1946, в-д - см. обозначения на рис. 30
Самые крупные растения, образующие довольно обширные заросли в сублиторали западной части Кандалакшского залива, вдоль Карельского и Поморского берегов, значительно крупнее изредка встречающихся сублиторальных (и самых крупных) растений Восточного Мурмана (рис. 33).
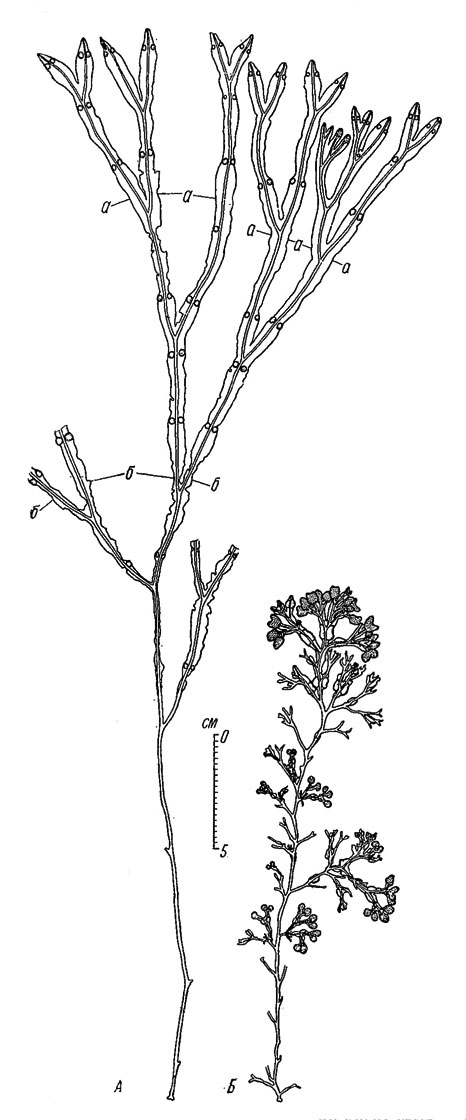
Рис. 33. Самые крупные растения Fucus vesiculosus. А - из Белого моря (Карельский берег, губа Грицина): а - нижняя граница живой части растения, б - нижняя граница отмирающей части растения; Б - с Восточного Мурмана (кут губы Ярнышной)
Анализ некоторых биологических свойств различных популяций F. vesiculosus в Баренцевом и Белом морях (табл. 28) позволяет сделать следующие краткие заключения:
- Максимальная продолжительность жизни растений в Баренцевом. (12 + ) и Белом (11 + ) морях практически одинакова*.
- Максимальная ширина "ветвей" слоевища в Белом море (31 мм) достигает большей величины, чем в Баренцевом (21 мм).
- Наибольший годовой прирост длины слоевища в Белом море (213 мм) значительно больше того, что наблюдается на Мурмане (184 мм). Причем в Баренцевом море указанный размер прироста бывает очень редко и у единичных растений; в массовых зарослях годовой прирост здесь не превышает 130 мм. В Белом море годовой прирост длины слоевища в 150-200 мм и больше можно считать обычным явлением, свойственным растениям массовых зарослей не только сублиторали, но и литорали.
- Среднее количество концептакул в одной рецептакуле у беломорских растений (381) больше, чем у мурманских (268).
- Максимальная биомасса растений на единицу площади на Восточном Мурмане, как уже говорилось ранее, достигает более высоких показателей, чем в Белом море.
* ( К. П. Гемп и З. Г. Паленичко (1956) сообщают, что в Белом море длительность жизни растений этого вида составляет 10-20 лет. В наших обширных сборах, охвативших все беломорские, побережья, растений со столь высокой длительностью жизни не обнаружено. Возможно, что если они и встречаются, то крайне редко.)
| Место наблюдений | Средний прирост в длину за год (в мм) | Ширина пластинки (в мм) | Максимальный возраст (годы) | Среднее количество концептакул в одной рецептакуле (двойной/одинарной) | Максимальная биомасса (в кг/м2) |
| Восточный Мурман | |||||
| Подвергающиеся резкому опреснению кутовые участки губ У верхней границы зарослей |
84 | 8 | 5+ | 98/95 | - |
| У нижней границы зарослей | 86 | 13 | 8+ | 224/158 | - |
| Единичные растения у границы отлива | 184 | 21 | 12+ | 220/137 | - |
| Не подвергающиеся сильному опреснению кутовые участки губ У верхней границы зарослей |
104 | 9 | 5+ | 139/97 | 18.1 |
| У нижней границы зарослей | 130 | 13 | 7+ | 268/156 | 49.9 |
| Каменистые россыпи открытых берегов У верхней границы зарослей |
38 | 6 | 3+ | - | 9.6 |
| У нижней границы зарослей | 64 | 10 | 3+ | - | 16.6 |
| Отвесные и ступенчатые скалы на открытых берегах В средней части зарослей |
26 | 4 | 3+ | - | 6.5 |
| Воронка Белого моря | |||||
| Кутовые участки губ У верхней границы зарослей |
44 | 4 | 6+ | 197/115 | - |
| У нижней границы зарослей | 95 | 7 | 3+ | 252/142 | - |
| Каменистые россыпи на полуоткрытых берегах У верхней границы зарослей |
85 | 7 | 4+ | 197/115 | - |
| У нижней границы зарослей | 102 | 8 | 6+ | 232/142 | 13.4 |
| Горло Белого моря | |||||
| Каменистые россыпи закрытых берегов В средней части зарослей |
56 | 8 | 6+ | 102/61 | 41.7 |
| Каменистые россыпи открытых берегов | 89 | 9 | 5+ | 192/104 | - |
| Бассейн Белого моря | |||||
| Закрытые участки южного берега У верхней границы зарослей |
81 | 7 | 5+ | 186/91 | 10.8 |
| У нижней границы отлива | 106 | 9 | 8+ | 192/107 | 13.0 |
| На глубинах 0.5-1 м | 211 | 31 | 10+1 | Размножается очень редко | - |
| Открытые участки южного берега У верхней границы зарослей |
52 | 7 | 4+ | 98/44 | 19.8 |
| У нижней границы отлива | 101 | 10 | 6+ | 146/89 | 24.0 |
| На глубинах 0.5-1 м | 202 | 23 | 9+ | 210/109 | - |
| Открытые участки северного берега У верхней границы зарослей |
52 | 6 | 3+ | 245|119 | - |
| У нижней границы отлива | 112 | 8 | 5+ | 187/117 | 14.9 |
| Онежский залив | |||||
| Закрытые участки Поморского берега У верхней границы зарослей |
70 | 8 | 7+ | - | - |
| У нижней границы отлива | 152 | 13 | 7+ | - | - |
| На глубинах 0.5-1 м | 151 | 20 | 5+ | 155/75 | - |
| Открытые участки Лямицкого берега У нижней границы отлива |
107 | 6 | 3+ | 212/105 | -3 |
| На глубинах 0.5-1 м | 185 | 7 | 5+ | 155/75 | - |
| Открытые берега островов южной части залива У верхней границы зарослей |
103 | 6 | 5+ | - | - |
| У нижней границы отлива | 213 | 14 | 6+ | 269/132 | - |
| Закрытые берега островов средней части залива У верхней границы зарослей |
103 | 7 | 5+ | 195/97 | - |
| У нижней границы отлива | 167 | 17 | 5+ | 254/122 | - |
| На глубинах 0.5-1 м | 181 | 20 | 4+ | 381/198 | - |
| Открытые берега островов средней части залива У верхней границы зарослей |
83 | 6 | 3+ | 93/55 | - |
| У нижней границы отлива | 142 | 12 | 7+ | 137/87 | - |
| На глубинах 0.5-1 м | 175 | 19 | 5+ | 207/112 | - |
| Открытые берега островов северной части залива У верхней границы зарослей |
109 | 12 | 2+ | 235/139 | - |
| У нижней границы отлива | 133 | 8 | 4+ | 196/115 | - |
| На глубинах 0.5-1 м и более | 179 | 11 | 6+ | 306/142 | - |
| Кандалакшский залив | |||||
| Устьевые участки губ У верхней границы зарослей |
93 | 9 | 6+ | - | 25.8 |
| У нижней границы отлива | 108 | 11 | 7+ | - | 26.5 |
| На глубинах 0.5-1 м | 151 | 18 | 9+ | - | - |
| Кутовые участки губ У верхней границы зарослей |
50 | 9 | 6+ | - | 7.5 |
| У нижней границы отлива | 96 | 13 | 9+ | - | 21.8 |
| На глубинах 0.5-1 м | 145 | 22 | 11+ | - | - |
В прибрежных водах Англии, по данным весьма обстоятельной работы Найт и Парк (Knight, Park, 1950), продолжительность жизни растений этого вида едва ли многим превышает 3 года, а длина слоевища достигает 1 м и более. Средний годовой прирост длины слоевища в разных местах у растений различных возрастных групп составляет там от 161 до 353 мм, т. е. максимальная величина прироста значительно превышает не только то, что наблюдается на Восточном Мурмане, но и в Белом море. Это становится понятным, если учесть, что в прибрежных водах Англии рост идет непрерывно в течение круглого года. Выше было уже указано, что на литорали открытых участков Восточного Мурмана нередко можно заметить образование рецептакул и их созревание в любое время года, обычно же в этом отношении и здесь соблюдалась строгая сезонность. В прибрежных водах Англии заложение рецептакул, как правило, растягивается на период с октября по февраль, зрелые рецептакулы имеются с февраля по май, а сбрасывание их протекает с мая по октябрь. Иными словами, в отличие от строго установившейся сезонности в росте и размножении основной массы растений этого вида на литорали Восточного Мурмана и в Белом море, на литорали Англии эта сезонность в отношении роста исчезает вовсе, а размножение растягивается на значительно более долгий период года. Одновременно с этим в наших водах значительно увеличивается продолжительность жизни.
В некоторых участках Балтийского моря отдельные биологические свойства F. vesiculosus занимают промежуточное положение между тем. что наблюдается у североморских растений, с одной стороны, и мурманских и беломорских, с другой, причем они ближе к беломорским, чем к мурманским. Известно, например, что в Финском заливе растения этого вида могут достигать длины 2-3 м и в этом отношении сходны с растениями из Онежского залива Белого моря (Colman, 1933).
Довольно детально жизненный цикл F. vesiculosus изучен в западной части Балтийского моря в районе острова Хиддензее (Bauch, 1955/56; Kalähne, Wartenberg, Bauch, 1955/56; Künzenbach, 1955/56; Müller-Stoll u. Kü nzenbach, 1956), результатами этого исследования мы и воспользуемся для сравнения. Прежде всего скажем несколько слов об. условиях, в которых находятся водоросли в районе острова Хиддензее. Средняя температура поверхности воды в марте составляет здесь 2, в мае 13, в августе 22 и в октябре 10°. Абсолютный минимум зимой достигает - 0.2, а абсолютный максимум летом от 25 до 28°. Ледяного покрова, как правило, не бывает. Характерны низкие солености: в среднем соленость воды составляет около 8%о, с колебаниями в течение года от 7.9 до 9.4.
Заросли F. vesiculosus располагаются здесь на глубине около 2 м. Вдоль открытых берегов растения достигают длины 1.2-1.5 м, а на закрытых участках побережья не более 0.3-0.5 м. Плавательные пузыри образуются одновременно с началом роста слоевища, в конце марта или, чаще всего, в начале апреля. Вслед за первым рядом могут образовываться дополнительные пузыри более мелких размеров: самое большее за сезон роста образуется 6 пар плавательных пузырей. Продолжительность жизни и скорость роста водоросли, так же как на Мурмане и в Белом море, легко определяется по плавательным пузырям. Средний годовой прирост длины слоевища составляет здесь 18 см, при колебаниях от 12 до 27 см. Годовой прирост более 22 см встречается очень редко. Из этого видно, что годовые приросты балтийских и беломорских растений весьма близки. Уже к сентябрю средний прирост длины слоевища равен 15.5 см, т. е. всего на 2.5 см меньше общего среднего прироста за весь период вегетации. Эти 2.5 см вырастают в течение сравнительно холодного времени года, с октября по январь. В январе и феврале рост слоевища, если и имеет место, то идет очень медленно и при обычных наблюдениях в природе не обнаруживается.
Растения с длиной слоевища 120-150 см имеют возраст 6-7 лет, но большая часть растений не живет более 4-5 лет. Таким образом, растения из западной части Балтийского моря отличаются от мурманских и беломорских несколько уменьшенной продолжительностью жизни, а от североморских - несколько повышенной.
Рецептакулы у балтийских растений начинают развиваться в январе-феврале, достигают максимального количества и зрелости в июле и разрушаются в августе. В отличие от Мурмана и Белого моря, сбрасывание органов плодоношения происходит очень дружно и обычно после штормовой погоды на берегу скопляется масса пожелтевших рецептакул. Можно напомнить, что в наших водах только у Ascophyllum nodosum сбрасывание рецептакул осуществляется в течение короткого периода и заканчивается сразу же после плодоношения; у всех других фукоидов, в том числе и у Fucus vesiculosus, это происходит в течение нескольких месяцев.
В западной части Балтийского моря у небольшой части растений, обитающих на сравнительно больших глубинах, в июле могут образовываться молодые незрелые репептакулы, которые заканчивают свое развитие осенью, т. е. и в этом отношении имеются некоторые общие черты с мурманскими и беломорскими растениями. Мы уже говорили о том, что в условиях наиболее сглаженных колебаний факторов внешней среды (на скалах открытых берегов Восточного Мурмана и в сублиторали некоторых участков Белого моря), кроме основного периода размножения, бывает еще дополнительный, охватывающий небольшую часть растений.
Наши небольшие наблюдения к западу от г. Лиепая показали, что наиболее крупные растения имеют здесь слоевище длиной 110-120 см и возраст 4-6 лет. Средний годовой прирост слоевища таких растений составлял 253 при колебаниях от 235 до 275 мм. В течение года в этих условиях образуется обычно один, редко два ряда плавательных пузырей. Пластинка слоевища была тонкой и легко рвалась. Характером окраски и слабой прочностью растения этой части Балтийского моря напоминали собой растения из наиболее прогреваемых участков южной и средней части Онежского залива Белого моря. Наши наблюдения были проведены 19 июля 1951 г. К этому времени все взрослые растения были снабжены хорошо развитыми и еще полностью сохранившимися репептакулами.
В северо-восточной части Балтийского моря, в районе островов Эрегрунд, еще встречаются значительные заросли F. vesiculosus. Растения здесь живут при солености 2-4‰ и при довольно суровых температурных условиях: в летние дни температура поверхности воды может достигать 20, средняя в августе составляет 14, а в мае 2-3°. Зимой температура воды часто бывает ниже нуля, и ледяной припай толщиной 25-30 см держится в течение 30-40 дней. Растения здесь обычно лишены плавательных пузырей и у верхней границы зарослей имеют сильно ветвящееся слоевище, высотой до 25-30 см. У нижней границы зарослей, на глубине около 9 м, размеры растений не превышают 10-15 см. Годовой прирост длины слоевища, судя по растениям, имеющим плавательные пузыри, колеблется от 3 до 10 см. На одном растении бывает не более двух рядов плавательных пузырей и лишь изредка имеется еще одна пара пузырей в месте первого ветвления слоевища. Формирование рецептакул начинается в январе (Waern, 1952).
В различных частях ареала время начала развития рецептакул изменяется в пределах от октября до января-февраля, а длительность их созревания колеблется от 3 до 8-10 месяцев (табл. 29).
| Место наблюдений | Месяцы | Длительность созревания (месяцы) |
Авторы | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |||
| Восточная Гренландия | + | * | * | * | 0 | 0 | - | - | * | * | * | + | 5 | Jonson, 1904 |
| Юго-западная Исландия | * | * | * | * | * | * | * | 0 | 0 | + | * | * | 10(?) | Jonson, 1912 |
| Южная Англия | + | +0 | 0 | 0 | 0- | - | - | - | - | +- | + | + | 4 | Knight, Park, 1950 |
| Побережье Голландии | * | * | * | 0 | 0 | - | * | * | * | + | + | * | 7 | Van Goor, 1923 |
| Западная часть Балтийского моря | + | + | * | 0 | 0 | 0 | 0- | - | * | * | * | + | 4 | Kalähne, 1955/56 Müller-Slroll, Künzenbach,1956 |
| Северо-восточная часть Балтийского моря |
+ | * | * | * | * | * | * | * | * | * | * | * | ? | Waern, 1952 |
| Юго-запад Скандинавии | * | * | * | * | 0 | * | * | * | * | * | + | * | 6 | Sundene, 1953 |
| Средняя Норвегия | + | + | * | 0 | 0 | 0 | 0 | * | * | * | * | * | 3 | Printz, 1926 |
| Восточный Мурман | +- | * | * | * | 0 | 0- | - | - | - | - | - | +- | 6 | Наши наблюдения |
| Белое море | + | * | * | * | * | 0- | 0- | - | - | - | - | + | 7-8 | Наши наблюдения |
Примечание. + заложение рецептакул, 0 зрелые рецетакулы, - сбрасывание рецептакул.
Весь рассмотренный материал свидетельствует о крайне высокой эврибионтности F. vesiculosus, способном жить в самых различных условиях температуры и солености и имеющем благодаря этому чрезвычайно широкое географическое распространение.
В заключение добавим еще сведения об относительном весе вегетативных и генеративных частей слоевища у растений в пору их зрелого плодоношения:
| * | Количество рецептакул |
Вес (в %) |
| Восточный Мурман Отвесные и ступенчатые скалы на открытых берегах. Сильный прибой. Источников опреснения нет |
3160 | 37.6 |
| Валунные и каменистые россыпи на открытых берегах. Прибой слегка ослаблен. Источников опреснения нет |
2763 | 41.2 |
| Отдельные камни на илисто-песчанистых пляжах закрытых берегов. Прибоя нет. Источников опреснения нет |
3270 | 57.3 |
| Отдельные камни на илисто-песчанистых пляжах закрытых берегов. Прибоя нет. Ежесуточные колебания солености: в отлив соленость падает до 5-6, а в прилив поднимается до 29-30 ‰ |
2078 | 41.2 |
| Южный берег Бассейна Белого моря (район мыса Кирбей) Каменистые россыпи на открытых берегах. Сильный прибой. Источников опреснения нет |
4242 | 61.3 |
| Каменистые россыпи на полузакрытых берегах. Прибой слабый. Источников опреснения нет |
2079 | 36.0 |
| Пологие скалы на закрытых берегах. Прибоя нет. Источников опреснения нет |
1784 | 13.0 |
| Отдельные камни на илисто-песчанистых пляжах закрытых берегов. Прибоя и источников опреснения нет |
1364 | 36.0 |
В Белом море вес генеративных частей слоевища, ежегодно сбрасываемых после периода размножения, достигает 61.3 от общего веса растений, а на Мурмане 57.3%, в большинстве же случаев относительный вес генеративных частей у беломорских растений меньше, чем у мурманских, что обусловлено, по-видимому, увеличением общих размеров слоевища в Белом море. Относительный вес генеративных частей слоевища показывает, что ежегодно в общий круговорот веществ водоема поступает от 13 до 61 % общей биомассы F. vesiculosus в Белом море и от 38 до 57% на Восточном Мурмане; в среднем можно принять размер этой "продукции" равным 40-45% летней биомассы.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'