Оценка численности и биомассы облавливаемых стад рыб
Оценка величины стад промысловых рыб возможна как в абсолютных, так и в относительных показателях. Оценка абсолютной величины стада рыбы обычно заключается в определении численности или биомассы рыб, находящихся на определенной площади или в целом водоеме. Обычно оценку абсолютной численности производят в отношении промысловой части стада или подрастающего пополнения отдельно, так как методики оценки взрослых рыб и молоди обычно несколько различаются.
Метод площадей
Наиболее старым, но до сих пор не потерявшим своего значения методом является так называемый метод площадей. Принцип его заключается в том, что количество добытой на определенной площади рыбы относится ко всей площади водоема или к площади, занятой данным скоплением. Этот метод впервые был применен Гензеном и Апштейном (1897), которые на основании учета пелагической икры попытались определить общую численность нерестующего стада. Зная количество выметанных икринок в единице объема воды, среднюю плодовитость самки и соотношение полов в нерестовом стаде, они вычислили величину стада нерестящихся в данном районе рыб.
Их способ расчета может быть представлен в следующем виде:
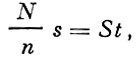 (30)
(30)где:
N - общее число выметанных икринок в обследованном районе;
n - средняя плодовитость самки;
s - соотношение полов в нерестовом стаде;
St - величина нерестового стада.
В свою очередь
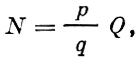 (31)
(31)где:
р - среднее число икринок в улове;
q - обловленный объем;
Q - общий объем воды в исследованном районе, где проводился учет.
Однако для правильного определения абсолютной численности нерестового стада этим способом требуется учет столь многих поправок, что практически этот метод оказывается применимым лишь в редких случаях. Погрешности метода связаны с недоучетом гибели развивающейся икры на разных стадиях, неравномерностью ее распределения в толще воды, недостатками в конструкции орудий лова, отражающимися на их уловистости. Т. С. Расе (1949) правильно указал, что, не зная коэффициента уловистости орудий лова в отношении икринок или, точнее, отношения числа выметанных икринок в единице объема к числу попавшихся в орудие лова, получить данные об абсолютной численности нерестового стада невозможно. Однако для получения относительных показателей по сравнению с предшествующими годами и с другими районами метод оценки относительной численности нерестового стада по количеству икринок в единице объема во время нереста вполне применим и в настоящее время.
Обзор работ о применении метода учета пелагической икры для оценки величины стада сделал Саундерс Инглиш (Saunders English, 1964). Этот автор считает, что рассматриваемым методом возможно оценивать колебания численности стада рыбы с пелагической икрой, особенно камбал. Были также сделаны попытки оценить численность нерестового стада рыб, откладывающих икру на дно. Так, Руннстрем (Riinnstrom, 1941) провел учет икры сельди Clupea harengus (L.) при помощи дночерпателя Петерсена у берегов Норвегии. Сопоставляя количество отложенной икры и уловы сельди в период нереста в этом районе, Руннстрем, кроме общей оценки величины нерестового стада, попытался определить и величину промысловой смертности сельди за весеннюю путину.
Стивенсон и Отрем (Stevenson a. Outram no Saville, 1964) на основе учета отложенной икры попытались определить величину нерестового стада и промысловой смертности сельди Британской Колумбии. Величины промысловой смертности этой сельди по возврату помеченных рыб за ряд лет оказались близкими.
Таким образом, метод учета стада рыбы по отложенной донной икре в ряде случаев также может быть использован.
Принципы оценки численности стада на основании учета на единицу площади с последующей экстраполяцией на весь район, занятый скоплением, нашли широкое применение в дальнейшем. Обзор этих методов дан Г. Н. Монастырским (1940, 1952) и С. В. Аверинцевым (1948).
Для оценки численности объектов тралового рыболовства обычно пользуются следующей формулой:
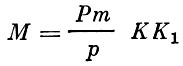 (32)
(32)где:
Р - площадь района, в котором проводится учет;
р - площадь, облавливаемая тралом в единицу времени;
m - средний улов в единицу времени;
К - коэффициент горизонтальной уловистости трала;
К1- коэффициент вертикальной уловистости.
При практическом применении указанного метода труднее всего определить коэффициенты К и K1 которые характеризуются видовой специфичностью и зависят от возраста и биологического состояния рыбы, времени суток, погоды и других моментов. Кроме того, селективность кутцов тралов зависит от плотности облавливаемой популяции и режима работы, что тоже приходится учитывать при оценке численности разных размерных групп рыб в облавливаемом скоплении.
Во многих случаях при оценке абсолютной численности стада методом площадей надежнее пользоваться кошельковыми орудиями лова. Принцип здесь остается тот же, что и при лове тралом, но точность учета многих рыб возрастает, так как при обработке данных лова на небольших глубинах, например в Азовском море (Майский, 1940; Смирнов, 1937), из формулы можно исключить коэффициент вертикальной уловистости, а коэффициент горизонтальной уловистости в отношении многих видов оказывается близким к 100%. Однако последний очень сильно зависит от соответствия величины орудия лова и облавливаемого скопления. Если орудие лова может обметать все скопление, то коэффициент горизонтальной уловистости обычно близок к 100%. Если же орудия лова захватывают лишь часть скопления, то уловистость резко снижается, так как рыба успевает уйти из зоны облова. Естественно, что величина коэффициента уловистости зависит также от состояния рыбы.
Дальнейшим развитием метода площадей применительно к пелагическим рыбам было сочетание обловов тем или иным орудием лова с определением площадей, занимаемых косяками, с самолета (Голенченко, 1950).
Качественный скачок в развитии метода площадей связан с использованием в рыбной промышленности гидролокационных приборов, позволяющих оконтуривать скопления рыб и по интенсивности записи скопления в сочетании с опытными обловами определять абсолютную численность рыб в скоплениях (Юданов, 1960).
Проведены интересные опыты по оценке абсолютной численности скоплений зимующей сельди путем сочетания эхометрических записей, при помощи которых определяют границы скопления и относительную величину концентрации рыб в разных местах скопления, и наблюдений телевизионной камеры, позволяющей определять число рыб в единице объема воды. Таким образом удается с практически достаточной точностью определить абсолютное количество рыб в скоплении. Опытные ловы служат в этом случае как бы контролем.
Комбинированный способ оценки численности скоплений сельди путем использования гидролокационных приборов, подводного телевидения и обловов разработан С. С. Федоровым, И. Д. Трускановым и И. Г. Юдановым (Fedorov, Truskanov a. Yudanov, 1964).
В настоящее время, пользуясь гидролокационными приборами, удается с достаточной точностью оценивать величину скопления и во многих случаях определять и основной вид рыб его образовавший (Love, 1971).
В небольших водоемах и изолированных бухтах абсолютный учет рыб возможен путем умерщвления всего рыбного населения теми или иными ядовитыми веществами. В этом отношении особый интерес представляют исследования Е. В. Бурмакина (1961, и др.), позволяющие определить общую биомассу рыбного населения озер различного типа и увязать их рыбопродуктивность с кормовой базой.
Несомненно, что в дальнейшем с совершенствованием гидролокационной техники и техники подводных наблюдений оценка методом площадей численности и биомассы стад промысловых рыб, образующих скопления и ведущих стайный образ жизни, будет приобретать все большее значение. Для оценки численности рыб, держащихся разреженно, метод площадей, видимо, будет иметь меньшее значение.
При оценке продуктивности водоемов по промысловым уловам нужно всегда учитывать, что лов ведется обычно в местах концентрации рыбы. Как отмечают Ю. Ю. Марти и Г. В. Мартинсен (1969), за одно притонение неводом при облове в дельте Волги полупроходных рыб во время хода может быть "снят урожай" с 3-5, а иногда и с 10-15 км2 Северного Каспия. Большой современный тральщик за час траления "снимает урожай" минимум с 1-2 км2, а за одно кошелькование современным кошельковым неводом часто "снимается урожай" с площади до 10 км2.
Методы учета численности движущихся рыб
Для учета численности стада проходных и полупроходных рыб существенное значение имеют способы оценки количества рыбы, прошедшей за определенный отрезок времени через поперечное сечение реки. Впервые у нас этот метод оценки численности мигрирующего стада был предложен Ф. И. Барановым (1933, 1960) для учета численности мигрирующей воблы. Для учета пользовались закидным неводом. Ф. И Баранов принимает, что общее количество рыбы, прошедшее через облавливаемый участок реки, равно:
где:
n - число рыб на единицу площади;
v - скорость хода рыбы;
Y - ширина реки в месте лова;
Т - время, прошедшее между последовательными заметами.
Однако таким способом лишь в редких случаях удается установить абсолютную численность прошедших рыб. Это связано с неравномерностью распределения движущихся рыб поперек русла, разной уловистостью невода в отношении рыб, встречающихся с неводом в разные фазы притонения. Этот метод более применим для учета относительной численности мигрирующих рыб.
Наиболее широко применяется количественный учет проходных дальневосточных лососей во время их нерестовой миграции. Таким образом удается в течение ряда лет учитывать численность отдельных локальных стад, заходящих для нереста в определенные реки. Многолетний материал накоплен сейчас в отношении ряда стад, например нерки Оnсоrhynchus nеrkа (Walb.) рек Озерной и Паратунки; кеты О. keta (Walb.) и горбуши О. gorbuscha (Walb.) реки Мы и некоторых других стад. Обычно просчет проводится визуально путем учета рыбы, периодически пропускаемой через специальные окошки в перегораживающих реку сплошных заграждениях.
На Аляске проходящих к нерестилищам рыб просчитывают путем периодической киносъемки рыб, пересекающих выложенные на дне белые панели, и путем экстраполяции пересчитывают на сутки (Mathisen, 1962 a). Затем, используя соответствующий эмпирический переводной коэффициент, подсчитывают абсолютное количество рыб, прошедших за определенный промежуток времени данное сечение реки. В 1955 г. в день снимали в среднем 1440 кадров.
Разница между данными, полученными путем непосредственного просчета и путем пересчета рыб с кинокадров, оказалась очень небольшой. В 1955 г. она составляла в среднем 1,91%.
В. Р. Протасов и Ю. А. Митрохин (1960) сконструировали прибор, позволяющий просчитывать количество и измерять длину рыб, проходящих через определенный участок (рис. 109). Рыбы, проходящие через определенный отсек, прерывают свет; тем самым отмечаются как длина рыбы, так и число рыб, прошедших через определенное отверстие. Недостаток метода заключается в том, что он не позволяет учитывать видовую специфику проходящих рыб; кроме того, в некоторых случаях две, проходящие непосредственно одна за другой рыбы могут сливаться. Несомненно, что в дальнейшем для количественного учета мигрирующих рыб автоматика будет использоваться значительно шире. Основная сложность заключается в обеспечении раздельного учета при совместной миграции нескольких видов рыб (Протасов и Неймарк, 1964).
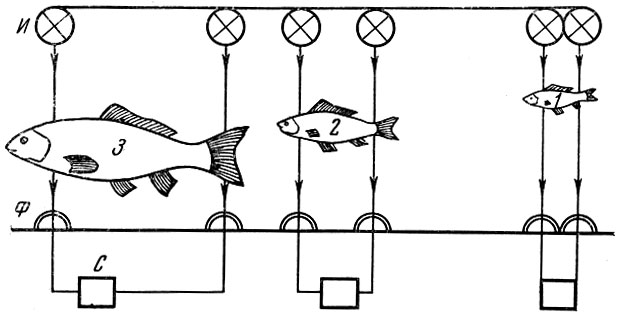
Рис. 109. Схема размещения фотоэлементов и источников света, позволяющая регистрировать рыб по размерам (по Протасову и Митрохину, 1960): Ф - фотоэлементы; И - источники света; С - счетчики; 1, 2, 3 - учитываемые размерные группы
Так же, как учет рыб при нерестовой миграции, в ряде случаев возможен и учет скатывающейся молоди проходных рыб, особенно молоди лососей. Редко, например на некоторых нерестовых ключах Камчатки, удается осуществить абсолютный учет всей скатывающейся молоди. Обычно же учет покатной молоди ловушками дает лишь относительные величины.
Учет численности стада путем мечения
Определение абсолютной численности стада рыбы при помощи мечения основано на допущении, что число помеченных рыб так относится к числу пойманных рыб с метками, как количество добытых промыслом рыб относится ко всему количеству рыб промыслового размера в водоеме, т. е. имеет место следующее соотношение:
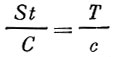 (34)
(34)где:
St - промысловое стадо;
С - величина вылова;
Т - число помеченных рыб;
с - число пойманных рыб с метками.
Однако практически, пользуясь этим соотношением, получить достоверные данные удается весьма редко; это возможно в отношении главным образом рыб с относительно длинным жизненным циклом и хорошо переносящих мечение (атлантический лосось, треска, лещ и др.).
Основные ошибки, связанные с применением этого метода, объясняются недоучетом поведения помеченных рыб, которые распределяются неравномерно среди остальных рыб стада. Часто поведение рыб с метками резко отличается от поведения непомеченных рыб (Державин, 1922; Караваев, 1939; Ricker, 1958; и др.). Процент потерянных меток рыбами, помеченными в разном биологическом состоянии, оказывается далеко не одинаковым, а это, естественно, влияет на результаты определения величины стада рыбы (Фортунатова и Чугунова, 1960). Неверный результат может получаться и потому, что смертность меченых рыб оказывается иной, чем немеченых (Parker, Black a. Larkin, 1963).
Большая смертность меченых рыб может объясняться тем, что рыбы, помеченные определенными типами меток, более интенсивно выедаются хищниками (Lawler and Smith, 1963). Как показал опыт, рыбы, вынутые из кутца, трала и помеченные на глубине под водой, выживают гораздо лучше, чем если они взяты из трала, поднятого на поверхность (Hislop and Hemmings, 1971).
Естественно, что смертность меченых рыб в очень большой степени зависит от того, в каком состоянии они были помечены (Beverton, Gulland'and Margets, 1959).
Определенная часть меток, найденная рыбаками, ими не возвращается. В Норвегии эта часть составляет 4-6% (Hylen, 1963).
Обычно ошибки в сторону занижения процента вылова получаются оттого, что в приведенной выше формуле учитывается мечение только рыб нерестовой популяции данного года, а в величину улова включается как остаток (от которого и надо вести расчет), так и пополнение, которое мечению не подвергалось.
Проверка точности определения численности путем мечения окуня Perca fluviatilis flavescens Mitch, в прудах, где могла быть учтена действительная численность рыбы, показала, что метод мечения в 54% случаев дал ошибку более чем 10% истинной величины популяции. В ряде же случаев ошибка была значительно большей (Buck, Toits, 1965).
В настоящее время метод мечения, как мне представляется, едва ли может иметь большое значение для оценки абсолютной численности стада. Его роль в анализе динамики популяций рыб сводится главным образом к определению некоторых показателей, в первую очередь таких, как смертность и выживание. Подробнее этот вопрос рассматривается в работах С. В. Аверинцева (1948), Г. Н. Монастырского (1952), Риккера (1958, и др.).
Оценка абсолютной численности стада рыбы по интенсивности выедания кормов
В некоторых случаях поголовье стада рыбы можно определить по интенсивности выедания ею кормов. Это определение, правда, относится обычно не к локальному стаду в целом, а его отдельным группировкам (Лебедев, 1955). Расчет осуществляется по следующей формуле:
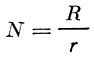 (35)
(35)где:
N - число кормящихся рыб;
R - общее количество корма, потребляемого всем стадом в единицу времени (рацион всего стада, учитываемый по непосредственным определениям динамики кормовой базы);
r - рацион одной рыбы.
Пользуясь этой методикой, часто удается примерно оценить величину кормящегося стада рыбы. Сложнее обстоит дело, когда приходится путем ловов устанавливать относительную численность каждого вида; зная средний рацион особи каждого вида, можно определить их численность, исходя из того же принципа, который заложен в приведенной выше формуле. В отношении рыб-бентофагов этот метод оказывается более применимым, чем в отношении рыб-планктонофагов.
Методы оценки относительной численности стада
Учет на основе анализа общих уловов и уловов на рыболовное усилие. После того как промысел достигнет определенной интенсивности, колебания уловов обычно отражают изменения численности промыслового стада. Следовательно, при более или менее стабильной интенсивности рыболовства по колебаниям уловов можно судить о колебаниях численности стада рыбы.
Колебания общего вылова (годовых уловов) обычно могут служить довольно надежным критерием изменения численности стада. Изменения уловов за более короткие промежутки времени часто могут отражать не столько изменения в численности и биомассе стада, сколько сдвиги в ходе годового цикла рыбы в связи с режимом данного года. Поскольку промысел обычно бывает приурочен к определенным фазам годового цикла (миграции, нерест, нагул и др.), то сдвиги этих сезонных явлений могут сильно отражаться на величине уловов за тот или иной отрезок времени.
Естественно, что оценка изменений численности и биомассы стада по колебаниям общих уловов требует учета и технико-экономических сторон рыболовства. Совершенствование техники рыболовства, естественно, приводит к относительному увеличению вылова.
Большое значение имеют и экономические причины: наличие спроса, себестоимость добычи и обработки рыбы, наличие рабочей силы и т. д. Все это, естественно, отражается на величине уловов.
Таким образом, колебания уловов обычно отражают изменения численности и биомассы стада рыбы при неизменной интенсивности рыболовства. Поскольку практически интенсивность рыболовства лишь очень редко остается стабильной, ее изменения приходится учитывать при анализе колебаний уловов.
Анализ общих уловов позволяет проследить как общую тенденцию изменения биомассы стада, так (при наличии раздельного учета рыб разного размера) и динамику примерного соотношения взрослого стада и пополнения. На основе анализа общих уловов удается выявить многолетние колебания величины стада и степень влияния рыболовства. Только хорошо налаженная промысловая статистика может быть использована для анализа динамики стада промысловых рыб. Хорошо налаженная статистика уловов - это показатель культуры рыболовства. Очень важно, чтобы группировка статистических данных соответствовала естественным внутривидовым группировкам промысловых рыб (например, летняя и осенняя кета, ивановская и егорьевская беломорская сельди и т. п.) и чтобы на основе анализа статистики уловов можно было выявить динамику уловов отдельных локальных стад. Хорошо налаженная статистика уловов обеспечивает более точную оценку рыбных ресурсов, более надежные прогнозы их изменений, а в конечном итоге все это значительно повышает эффективность рыболовства.
Основным показателем, наиболее широко используемым для оценки состояния стада и при составлении прогнозов колебаний численности и биомассы промысловых рыб, является улов на рыболовное усилие. Под уловом на рыболовное усилие понимается величина улова тем или иным орудием лова или кораблем за единицу времени. Характер применяемого показателя улова на рыболовное усилие зависит от специфики промышляемой рыбы и от применяемых орудий рыболовства.
Наименее надежным, сильно зависящим от ряда привходящих обстоятельств является улов на выгрузку, или улов на рейс судна. Обычно в научно-промысловых исследованиях используются улов на замет кошелькового невода (catch per shot), улов на сетку/дрейф при лове дрифтерными сетями, улов на ящик ярусной снасти за постановку (catch per scate) и т. п. Самым распространенным показателем улова на рыболовное усилие при анализе динамики стада донных рыб является улов на час траления. Этот показатель, пожалуй, лучше всего отражает изменения в состоянии численности стада исследуемой рыбы. Для того чтобы улов на рыболовное усилие отражал действительные изменения, происходящие в численности и биомассе стада рыбы, необходимо, кроме достаточного количества наблюдений, позволяющих получать статистически достоверные результаты, учитывать ряд моментов, связанных как с состоянием и поведением рыбы, так и с состоянием погоды и с техническими условиями лова. Естественно, что поскольку, например, уловистость трала, как и многих других орудий, меняется в отношении многих рыб в разное время суток, то и это приходится учитывать при анализе. Существенное значение имеют технические и конструктивные свойства орудий лова (окраска дрифтерных сетей, качество поводцов у ярусов и т. п.).
При анализе уловов на час траления очень важное значение имеют скорость траления и величина трала, что прямо связано с техническими показателями корабля и судового двигателя (Lundbeck, 1964).
Делались попытки использовать анализ уловов на рыболовное усилие и для оценки абсолютной численности стада. Эту цель, в частности, преследует регрессионный метод Де Лури (Rounsefell a. Everhart, 1953; Ивлев, 1958). Однако пользоваться этим методом для оценки абсолютной численности стада можно лишь в редких случаях, когда вылов значительно снижает численность популяции. Кроме того, этот метод предусматривает отсутствие пополнения стада в период промыслового сезона. Наконец, метод Де Лури требует наличия линейной корреляции между уловом на рыболовное усилие и общим уловом за время промысла, что, как известно, наблюдается далеко не всегда. Поэтому метод Де Лури применим, да и то с большими оговорками, лишь в очень редких случаях (о методе Де Лури см. Ивлев, 1958). Мне представляется также, что метод Де Лури слишком схематизирует биологические процессы, протекающие в водоеме.
Но если определение абсолютной численности путем анализа уловов на рыболовное усилие, видимо, имеет весьма слабые перспективы, то анализ этого показателя для оценки относительной численности стада заслуживает самого пристального внимания. Подробный обзор применимости показателя улова н'а рыболовное усилие сделан Гулландом (Gulland, 1964), и мы на деталях этого вопроса позволим себе не останавливаться.
Величина уловов на рыболовное усилие зависит от качества работы службы краткосрочных прогнозов (прогнозов миграций, распределения и концентраций) и оперативной разведки. Для таких показателей, как улов на выгрузку (catch per landing), улов на рейсосутки и т. п., хорошая работа указанной службы часто имеет решающее значение, сокращая до минимума холостые пробеги и затрату времени на поиск скоплений рыбы. Не меньшее значение при лове отцеживающими орудиями, в первую очередь тралом, имеет поисковая техника. Введение в рыбную промышленность эхолотов, особенно введение прицельного тралирования, естественно, весьма изменило величину уловов на рыболовное усилие. Часто при снижении численности стада удается некоторое время путем совершенствования техники поиска и лова поддерживать уловы на рыболовное усилие на высоком уровне. Указанное обстоятельство приходится учитывать при анализе динамики уловов на рыболовное усилие за длительные сроки. Пользоваться в этом случае различными поправочными коэффициентами следует очень осторожно, ибо иногда ничтожное, казалось бы, изменение в конструкции орудия или техники лова может коренным образом изменить уловистость. Это, конечно, не означает, что введение в ряде случаев соответствующих поправочных коэффициентов, полученных на основе биологически и статистически достоверных эмпирических сравнительных данных, невозможно. Такие коэффициенты вполне допустимы и широко применяются, например, при сравнении селективности и уловистости тралов, сделанных из разных материалов, или сетей разной окраски. Правда, в последнем случае уловистость резко меняется, как уже говорилось, в зависимости от освещенности, мутности, цвета воды и ряда других факторов.
Очень существенными критериями для суждения о динамике относительной численности стада часто может служить сопоставление таких показателей, как улов на рыболовное усилие, интенсивность рыболовства и величина общего вылова. На основе сопоставления этих показателей удается получить представление об изменениях численности и биомассы стада лучше, чем при анализе этих показателей изолированно.
Стандартизация учета рыболовного усилия в международном рыболовстве крайне важна для разработки согласованных мероприятий по ведению рыбного хозяйства в международных водах (Studenetsky, 1972).
Учет на основе анализа уловов и возрастного состава стада. Если допустить, что естественная смертность остается более или менее стабильной из года в год, то на основе анализа возрастного состава стада и общих уловов за ряд лет, равных по числу продолжительности жизни рыбы с момента вступления в промысловое стадо и до достигаемого рыбой предельного возраста, можно определить абсолютную численность выловленных рыб данного поколения. Сопоставляя рассчитанные подобным образом данные о численности отдельных поколений, можно составить представление о динамике численности стада за прошлый период и оценить изменения, происходящие в состоянии стада. Мы не останавливаемся на различных модификациях этого метода. Подробный разбор методик оценки относительной численности стада промысловых рыб на основе анализа возрастного состава стада дан Г. Н. Монастырским (1952) и Т. Ф. Дементьевой (1964).
Таковы те основные эмпирические приемы, которыми в настоящее время приходится пользоваться для оценки как абсолютной, так и относительной численности стад промысловых рыб. Несомненно, что в дальнейшем эти приемы будут совершенствоваться, причем основное направление развития методов оценки абсолютной и относительной численности стад рыб - это сочетание анализа уловов на рыболовное усилие с гидроакустическими и подводными телевизионными наблюдениями.
Однако рыбной промышленности нужно знать состояние сырьевой базы не только на сегодняшний день. Не менее важен прогноз предстоящих изменений сырьевой базы.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'