Модели с непрерывным временем
Впервые, насколько мне известно, дифференциальные уравнения были применены к анализу динамики популяций Россом (Ross, 1910). Дифференциальное уравнение "борьбы за существование между малярийным плазмодием и человеком", по Россу, имеет следующий вид (мы даем уравнения Росса с обозначениями, принятыми Г. Ф. Гаузе, 1933):
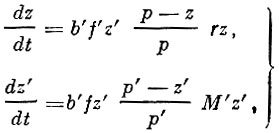 (12)
(12)где:
р - общее число людей в районе;
р'- число комаров в том же районе;
z - число людей, зараженных малярией;
z'- число комаров, содержащих плазмодии;
fz' - число заразных маляриков;
f'z'- число заразных комаров (fz' и f'z' составляют соответственно часть от z и z');
b'- количество укусов в единицу времени;
r - скорость выздоровления;
М' - смертность комаров;
t - время (см. также Гаузе, 1933).
Ряд положений Росса, в первую очередь принятие средней "мгновенной скорости заражения"
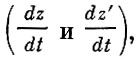
вошли и в построения Ф. И. Баранова (1918), который исходил из принципов, принятых в модели В. А. Кевдина (1915).
Ф. И. Баранов принял, что количество молоди не ограничивается величиной нерестового стада, т. е. молоди в водоеме всегда достаточно. Величина пополнения определяется количеством кормов, которое в водоеме постоянно и меняется только в результате воздействия стада рыбы. Неселективный промысел сам по себе может менять возрастную структуру облавливаемой популяции. Согласно Ф. И. Баранову, по изменению возрастного состава можно непосредственно судить об интенсивности смертности, а поскольку естественная смертность принимается за постоянную величину, то все изменения могут быть отнесены к действию вылова. В итоге своих исследований воблы Ф. И. Баранов (1925) выводит следующую математическую зависимость:
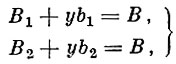 (13)
(13)где:
В1, В2 - ОСНОВНОЙ запас;
b1, b2 - вылов;
В - естественный запас;
γ - коэффициент.
Имея данные об уловах за ряд лет, Ф. И. Баранов пытается вычислить коэффициент γ, который он принимает постоянным, что, однако, не соответствует действительному положению дел. Баранов исходит из положения, что уменьшение числа рыб каждой возрастной группы за малый промежуток пропорционально численности этой группы и равно:
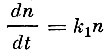 (14)
(14)или
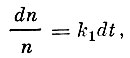 (15)
(15)где:
n - численность любой группы рыб;
t - время;
k1- коэффициент, одинаковый для всех групп.
Если известен возрастной состав улова, то численность стада есть сумма отдельных возрастных групп, которая может быть изображена в виде бесконечно убывающей геометрической прогрессии, т. е.
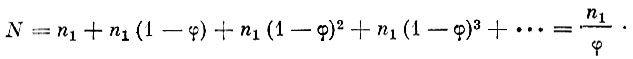 (16)
(16)При этом
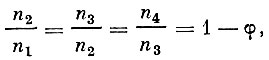
где: φ - относительная годичная убыль рыбы, равная
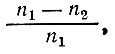
т. е. относительная величина убыли принимается как постоянная величина.
В своих представлениях Ф. И. Баранов, кроме точки зрения Кевдина (имя которого Баранов, к сожалению, не упоминал), в значительной мере использовал представления и материалы Петерсена (Petersen, 1895) и Гейнке (Heincke, 1913) по камбале Северного моря.
Ф. И. Баранов был первым исследователем, использовавшим дифференциальные уравнения для оценки динамики численности рыб. В дальнейшем дифференциальными уравнениями пользовались для моделирования динамики популяций рыб Бевертон и Холт (1957), рабочая группа но арктической треске (1959), Гулланд (Gulland, 1964) и многие другие исследователи. В общем виде современная модель динамики популяции, базирующаяся на дифференциальных уравнениях, может быть выражена следующим уравнением (Schaefer a. Beverton, 1963):
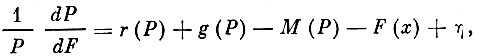
где:
Р - биомасса промысловой части популяции;
r - интенсивность пополнения;
g - темп роста;
М- интенсивность естественной смертности;
F - интенсивность промысловой смертности;
х - промысловое усилие;
η - показатель изменений популяции в зависимости от внешних условий вне связи с величиной популяции и интенсивностью рыболовства.
Большинство изменений, внесенных в последнее время в эту схему, не носит принципиального характера, а связано с фактическими сведениями и спецификой рыболовства. Так, модель Бевертона и Холта (1957) основана на результатах промысла донных рыб, а модель Шефера (1954) - на данных пелагического рыболовства. Принципиальной же разницы в них нет. (Подробнее о моделях этого типа см. Schaefer a. Beverton, 1963). Однако, как правильно отметил В. В. Меншуткин (1964), "биологическая емкость" моделей, использующих дифференциальные уравнения и исходящих из принципа непрерывного времени, значительно меньше, чем структурных моделей, основанных на принципе дискретного времени.
Стремление использовать хорошо разработанную теорию линейных дифференциальных уравнений для моделирования биологических процессов, в том числе динамики популяций животных, часто приводит к тому, что полученный результат не может быть интерпретирован биологически или результаты не вносят ничего нового в понимание биологического явления (Меншуткин, 1971). Модели с дискретным временем, использующие аппарат уравнений в конечных разностях, видимо в первую очередь и должны быть положены в основу при использовании ЭВМ.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'