Связь плодовитости родительского стада и численности потомства
Смертность особей, вызываемая теми или иными причинами, может очень сильно варьировать. Поэтому часто прямая связь численности родительского стада и потомства, опосредуясь в ряде звеньев, иногда сильно нарушается. Несомненно, что в сходных условиях большее количество производителей и выметанной высококачественной икры дают более многочисленное и более жизнестойкое потомство. Однако в природе из-за изменчивости условий размножения, развития проста поколений прямая корреляции числа отложенных икринок и величины пополнения часто нарушается.
У рыб с малой плодовитостью корреляция между числом отложенных икринок и пополнением обычно больше, чем у высокоплодовитых. По плодовитости, изменчивости и качеству запаса желтка в икре можно судить о состоянии нерестовой популяции, но только по этим показателям обычно нельзя давать промысловый прогноз будущего урожая. Мощность и качество пополнения нерестового стада - это результат взаимодействия количества и свойств отложенной оплодотворенной икры, определяющихся величиной и качеством нерестового стада, и тех абиотических и биотических условий, в которых происходят нерест, дальнейшее развитие икры,, рост и созревание пополнения.
Хотя количественная связь родительского стада и потомства часто и сильно вуалируется последующими условиями развития, все же в большинстве случаев удается установить более или менее выраженную связь изменений численности родительского стада с численностью продуцированного потомства.
Риккер (1954) прав, когда пишет: Взгляды по этому поводу (количественной связи родительского стада и потомства. - Г. Н.) колеблются между предположением о прямой пропорциональности зависимости между величиной взрослого стада и численностью пополнения и утверждением того, что численность пополнения фактически не зависит от величины взрослого стада.
Значительно реже затрагивается вопрос о возможности уменьшения численности пополнения при более высокой плотности родительского стада".
По этому вопросу ведутся довольно широкие споры. Встречаются такие, например, противоречивые высказывания.
"Для большинства рыб, обитающих в крупных водоемах, видимо характерно отсутствие прямой зависимости между величинами поколения и количеством производителей" (Танасийчук, 1957); "...связи между количеством икры и личинок рыб с последующей численностью поколений нет" (Ижевский, 1961); у "... трески и сельди все высокоурожайные поколения появились в периоды высокой производительной способности нерестовых популяций" (Марти, 1961); "... имеется хорошо выраженная зависимость между количеством прошедших на нерест производителей красной, скатившейся в море молоди и численностью взрослых рыб производного поколения" (Семко, 1961).
Исследователи, отрицающие закономерную связь между количеством икры, выметываемой родительским стадом, и численностью потомства, объясняют это явление по-разному. Одни (Баранов, 1918, 1925, и др.) исходят из представления, что молоди всегда достаточно, но в результате недостатка пищи может выжить лишь определенное количество. Другие (Дементьева, 1964; Ижевский, 1961, 1964, и др.), отрицающие связь плодовитости родительского стада и численности потомства, исходят из того, что условия жизни крайне изменчивы; поэтому от отложенного числа яиц выживает столь различное количество потомков, что закономерной связи родители - потомство проследить не удается.
Отрицание закономерной связи плодовитости и качественных показателей родительского стада с количеством воспроизводимого потомства приводит, естественно, и к определенным практическим выводам в области как эксплуатации стад промысловых рыб, так и их воспроизводства.
Как же обстоит дело в действительности? В отношении многих рыб показано, что между плодовитостью родительского стада и численностью потомства существует определенная количественная связь. Но эта связь, конечно, зависит от условий жизни и численности популяции. Несомненно, как это показал Р. С. Семко (1961), производительность популяции закономерно меняется при разных уровнях ее численности и обеспеченности популяции пищей. У дальневосточных лососей положительная корреляция выражена при определенных уровнях численности родительского стада: когда численность родительского стада превышает определенную величину, эта корреляция переходит в обратную (рис. 89). Эту связь у тихоокеанских лососей Томпсон (1962) выражает в виде теоретической кривой (рис. 90), которая оказывается близкой к эмпирической кривой, приводимой Семко (1961). Однако отмечаемая Р. С. Семко (1961) и Томпсоном (1962) закономерность свойственна далеко не всем стадам лососей, и если левая восходящая часть кривой прослеживается у всех стад, то правая нисходящая может отсутствовать, либо кривая после перегиба идет параллельно горизонтальной оси координат.
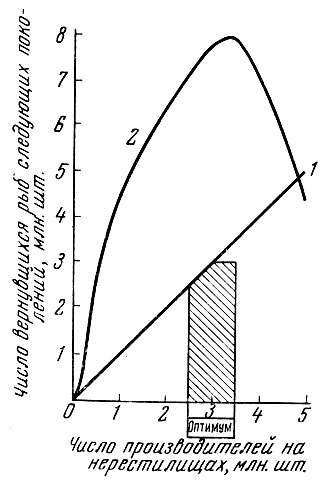
Рис. 89. Соотношение между числом производителей красной р. Озерной и вернувшихся рыб следующих поколений (по Семко, 1961): 1 - возвраты, равные числу производителей; 2 - фактические возвраты. Заштриховано необходимое количество рыб, пропускаемых для нереста
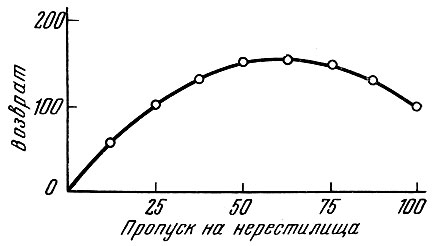
Рис. 90. Зависимость между числом производителей и величиной возврата у тихоокеанских лососей рода Oncorhynchus (no Thompson, 1962)
Попытаемся разобраться, каковы же количественные закономерности, которым подчиняются взаимосвязи родительского стада и потомства. При сходных условиях выживания численность потомства прямо пропорциональна плодовитости родительского стада.
В отношении осетровых Каспия Н. Л. Чугунов (1928) установил определенную связь между количеством молоди, скатившейся в море, и количеством производителей, пропущенных на нерестилища. Так, при запуске рыболовства в 1918 г. уловы осетровых упали по сравнению со средней многолетней почти в 3,5 раза; средний улов молоди осетровых на трал увеличился с 4,04 до 14,58 шт., т. е. в 3,6 раза. Связь между количеством производителей осетровых, пропущенных на нерестилища, и величиной возврата отмечена Н. Л. Чугуновым (1927) и у осетровых Азовского моря. Осетровые - рыбы со слабо выраженными флюктуациями численности, но и у рыб, у которых флюктуации выражены очень сильно, например у сельди, тоже наблюдается закономерная связь численности родительского стада и потомства. Как отмечает Ю. Ю. Марти (196J.), мощное поколение, как правило, воспроизводится от мощного поколения. По данным Иохансена (Johansen, 1930), имеется прямая связь между уловами нерестовой сельди и числом личинок (табл. 97).

Таблица 97. Промысловые уловы сельди в зунде и южной балтике и количество личинок сельди в 30-минутных ловах рингтралом (по Johansen, 1930)
У весенней салаки в районе Рюгена также отмечена связь численности нерестового стада и численности выведшихся личинок (табл. 98).

Таблица 98. Уловы весенней салаки в районе Рюгена и численность ее личинок (по Anwand a. Waldmann, 1962)
Как показывают наблюдения этих авторов, указанная связь прослеживается и в численности дочерних поколений, вступивших в нерестовое стадо в 1960 и 1961 гг.
Очень четкая связь имеется между численностью нерестового стада каспийских проходных сельдей и урожаем потомства, учтенным по ловам молоди в море (рис. 91).
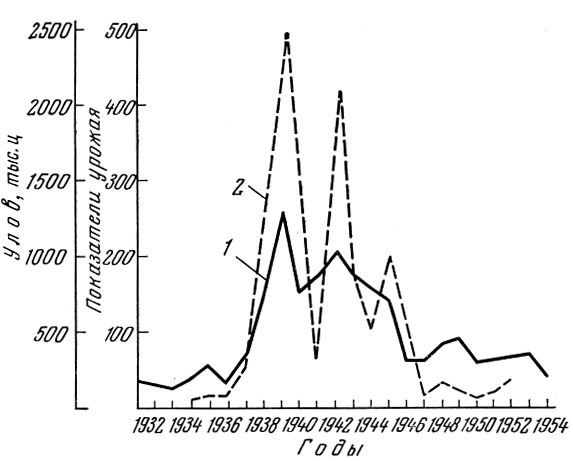
Рис. 91. Уловы сельди в Волго-Каспийском районе и показатели урожая пузанка и проходных сельдей 500 (по В. С. Танасийчук, 1957): 1 - уловы сельди (в тыс. ц); 2 - показатели урожая (в шт. на час траления)
График связи числа отложенных икринок и численности вступившего в промысел поколения черноморской хамсы показан на рис. 92. Если откинуть 1952 и 1959 гг., расположенные в пределах правого плеча построенной нами кривой, когда при высокой плодовитости поколения оказались очень малыми, то корреляция между численностью поколения и количеством отложенной икры оказывается весьма высокой (коэффициент корреляции +0,82), т. е. в семи случаях, располагающихся в пределах левого плеча кривой, из девяти проанализированных обнаруживается четкая прямая связь численности родительского стада и потомства. На графике, приводимом Р. М. Павловской (1963), очень хорошо видно, что корреляция между численностью родительского стада хамсы и потомства более четко выражена при средних значениях численностей родительского стада и менее четко при максимальных и минимальных значениях.
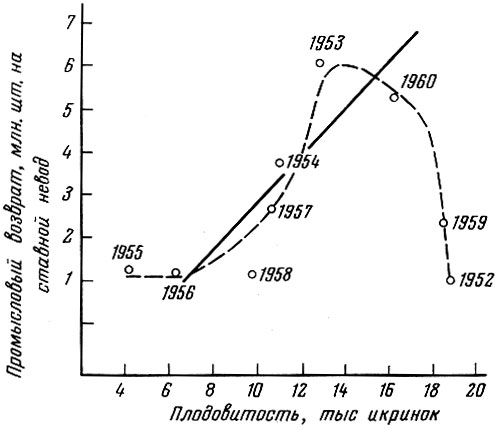
Рис. 92. Величина промыслового возврата поколений и средняя популяционная плодовитость черноморской хамсы (по Павловской, 1963) (кривая проведена нами)
Так как обеспеченность пищей обычно повышается одновременно и для родительского стада и для молоди, то увеличение плодовитости популяции обычно приводит к соответствующему увеличению численности потомства. Перегиб "кривой воспроизводства" имеет место тогда, когда условия жизни родительского стада меняются не адекватно условиям жизни потомства и приспособительная реакция родительского стада оказывается не соответствующей условиям выживания молоди.
Столь же четкая картина, очень напоминающая только что рассмотренную у хамсы, отмечена у дальневосточных лососей как Тихоокеанского побережья Америки, так и Азии (Ricker, 1954, и др.; Thompson, 1962; Семко, 1961, и др.). У лососей бассейна Амура установлены положительные коэффициенты корреляции между числом отнерестовавших производителей и количеством скатившихся из реки мальков. Так, по данным В. Я. Леванидова, у стад горбуши разных рек коэффициент корреляции колеблется от +0,59 ±0,28 (река Мы) до +0,84±0,33 (река Ул); у летней кеты - от 0,54±0,29 до +0,74±0,22, а у осенней кеты он равен +0,69±0,26.
В. Я. Леванидов (1964, 1969) считает, что в настоящее время плотность производителей лососей Амура на нерестилищах не достигает величины, которая бы отрицательно сказывалась на выживании икры. В то же время переполнение производителями нерестилищ приводит к расширению нерестовых площадей, их мелиорации путем перекапывания и к повышению кормности нерестовых водоемов для молоди за счет трупов родителей. Отрицательное влияние переуплотнения нерестилищ может сказываться через перекапывание нерестовых бугров позднее подошедшими производителями. Что же касается данных по ухудшению режима нерестовых водоемов и в частности условий инкубации икры в буграх, то их пока нет.
При численности стада лососей меньше максимальной "емкости реки" наблюдается четкая корреляция между плодовитостью родительского стада и численностью потомства. Если же в результате благоприятных условий, создавшихся в море, плодовитость стада превысит пропускную способность реки, то численность стада снизится или стабилизируется на определенном уровне. Причины, определяющие емкость реки, могут быть различными. Это и ограниченность нерестовых площадей, приводящая.при массовых заходах к перекопке нерестовых бугров (дальневосточные лососи), и недостаток убежищ от хищников (атлантический лосось рек Канады), и недостаточная обеспеченность молоди пищей (лосось некоторых рек Балтики, данные М. Н. Лишева, и форели Англии, Le Cren, 1961).
Пестрятка захватывает себе необходимую охотничью территорию, видимо, различную по площади при разной кормности реки, а особи, которые по тем или иным причинам не смогли получить себе необходимой охотничьей территории, уходят вверх, а чаще вниз по течению и в большей степени выедаются хищниками и гибнут от других причин. Если "кормовая емкость" реки недоиспользуется, имеется определенная корреляция между родительским стадом и потомством; если же "кормовая емкость" реки используется полностью, то ею в первую очередь и определяется численность стада лосося и стада форели того или иного речного бассейна.
У атлантического лосося Канады (Elson, 1962) до определенного предела имеется прямая связь между численностью родительского стада и численностью воспроизводящегося в данной реке потомства. Когда же количество молоди достигнет определенной величины, то далее ее численность гпределяется в первую очередь количеством укрытий от хищников, особенно рыбоядных птиц. Дальнейшее увеличение численности родительского стада уже не приводит к увеличению численности потомства, которая стабилизируется на определенной величине.
Ю. Е. Лапин (1971) на примере нерки Тихоокеанского побережья Америки отмечает прямую связь численности родительского стада и потомства и в то же время обращает внимание на то, что сначала рост численности родительского стада связан с увеличением колебаний урожайности потомства. Однако при максимальных величинах родительского стада амплитуда колебаний уменьшается, т. е. воспроизводство стада становится более устойчивым. Во всяком случае в определенных пределах и у лососей существует прямая связь между плодовитостью родительского стада и численностью потомства.
Прямая связь численности родительского стада и потомства отмечена и у некоторых популяций окуня оз. Байкал (рис. 93), например в заливе Мухор. Однако при благоприятных условиях нагула родительского стада (например, в Посольском соре Байкала) и плохих условиях выживания молоди имеет место закономерная обратная связь. Чем выше численность родительского стада, тем ниже (естественно, в определенных пределах) численность дочернего поколения ( рис. 93).
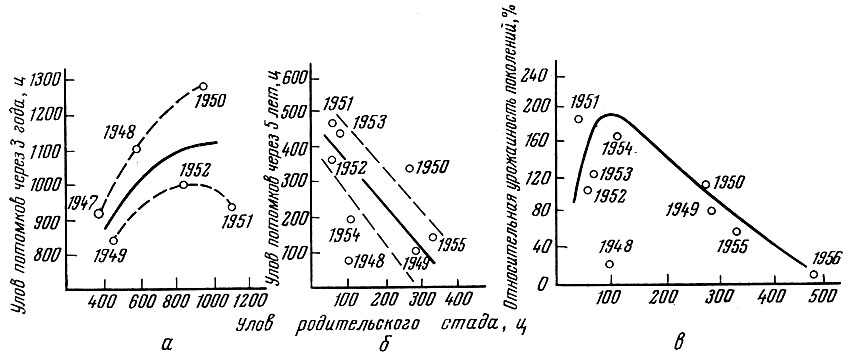
Рис. 93. Зависимость между численностью родительского стада и численностью взрослого потомства (по величине уловов) у разных популяций окуня оз. Байкал: а - в заливе Мухор; б - в Посольском соре; в - относительная урожайность поколений окуня Посольского сора, вычисленная по данным об уловах и возрастном составе за ряд лет и выраженная в процентах от среднего значения пополнения (по данным Б. К. Евтюховой). Цифрами на кривых обозначены годы рождения поколений
Весьма сходную характерную картину связи родители - потомство обнаружил Херрингтон (Herrington, 1948) у пикши Melanogrammus aeglefinus (L.) банки Джорджес (рис. 94). Интересно, что разброс точек на графике оказывается большим при минимальных и максимальных численностях родительского стада. Наличие закономерной связи численности родительского стада и потомства для аркто-норвежского стада трески отмечает Гаррод (Garrod, 1967).
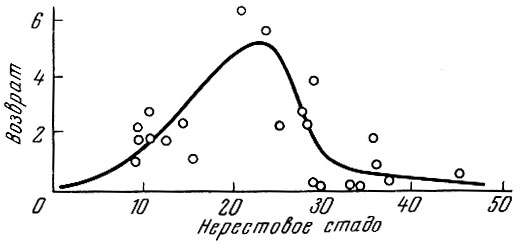
Рис. 94. Корреляция между родительским стадом и величиной потомства у пикши Melanogrammus aeglefinus (L.) банки Джорд-жес за 1914-1940 гг. (по Herring-ton, 1948). По оси абсцисс дана величина родительского стада (в тыс. англ. фунтов, на день промысла); по оси ординат - количество потомстяа (в тыс. шт. на день промысла)
Проведенные мною на рис. 89 и 92 прямые линии отражают тот случай, когда численность дочерней популяции при любых плотностях на всех этапах онтогенеза не оказывает влияния на выживание и рост пополнения. Однако, как отмечает Херрингтон (Herrington, 1948), в природе подобные случаи практически неизвестны. Выше мы показали, что у очень многих рыб обеспеченность внешней пищей на ранних этапах онтогенеза имеет важнейшее значение для выживания поколения. Чем дальше вправо сдвинута вершина кривой взаимоотношения численности родительского стада и потомства, тем выше обеспеченность пищей пополнения, т. е. темп воспроизводства меняется в соответствии с обеспеченностью пищей.
Поскольку популяция любого вида может регулировать интенсивность воспроизводства изменением плодовитости, то естественно, что связь между плодовитостью родительского стада и численностью потомства обычно нарушается тогда, когда условия жизни пополнения изменяются в противоположную сторону по сравнению с условиями, в которых жило родительское стадо.
В настоящее время у многих интенсивно облавливаемых видов рыб создалось такое положение, что плотность родительского стада редко достигает величины, при которой корреляция численности родительского стада и потомства становится отрицательной. Реализация правой части кривой соотношения родительского стада и потомства обычно имеет место в тех случаях, когда благоприятные условия, обеспечившие повышенную плодовитость стада, меняются в сторону ухудшения, что и приводит к повышенной гибели поколения.
В зависимости от того, какова численность родительского стада, может получиться как положительная корреляция (если точки находятся в левой части кривой), так и отрицательная (если точки расположены в правой части кривой). Отрицание связи между численностью родительского стада и потомства обычно и основывается на низких величинах коэффициентов корреляции (см. данные Павловской, 1963). Конечно, в ряде случаев не удается наблюдать корреляции между численностью потомства и численностью родителей даже в пределах только правого или левого "плеча" теоретической кривой.
Во многих случаях отсутствие прямой корреляции между числом выметанных икринок (или числом нерестовавших самок) и величиной урожая молоди в значительной мере объясняется тем, что не учитывается качество отложенной икры. Если бы сравнение велось с учетом качества половых продуктов, то корреляция, несомненно, получилась бы значительно большая. Ведь хорошо известно, что урожайность поколения определяется количеством и качеством выметанных родительским стадом половых продуктов и теми биотическими (пища, враги, болезни) и абиотическими (в первую очередь кислородный режим) условиями, в которых протекает развитие.
В. Е. Риккером (1954 и др.) много сделано для выявления закономерностей, определяющих количественную взаимосвязь численности родительского стада и воспроизводимого этим стадом потомства. Однако, мне думается, что Риккер недоучитывает значения качества отложенной родительским стадом икры в формировании дочернего поколения. Для отражения действительно существующей в природе взаимосвязи кривые воспроизводства, видимо, следует строить в системе трех координат: количество родителей, качество их половых продуктов и количество потомства. Фактически мы имеем дело с тремя, а вернее с четырьмя переменными: количество и качество потомства определяется количеством и качеством половых продуктов (как икры, так и молок!), продуцируемых родителями.
Под качеством мы понимаем соответствие условиям жизни, ибо то, что полезно в одних условиях, например разнокачественность икры, в других может оказаться вредным.
Недостаток построений Риккера заключается также в том, что количественные взаимосвязи родительского стада и пополнения он принял существующими как бы вне взаимосвязей со средой. Между тем если даже отбросить систему случайных связей, то и тогда взаимоотношения, складывающиеся в системе триотрофа, приведут к тому, что при разной напряженности этих отношений будут разные уровни воспроизводства, значит, мы будем иметь семейства кривых воспроизводства для разных популяций одного и того же вида, а в некоторых случаях уровни воспроизводства будут меняться и в пределах одной популяции, как это происходит у отдельных популяций дальневосточных лососей, например горбуши и летней кеты Амура, или у воблы и леща Северного Каспия. Следовательно, при разных уровнях напряженности вертикальных связей зависимость стадо - пополнение будет различной. Несмотря на эти оговорки, данные табл. 99 показывают, что существует видовая специфичность количества особей (в %), воспроизводимых от определенного стада и доживших до половозрелого состояния, так называемый "коэффициент промыслового "возврата". Это значит, что существует определенная связь между плодовитостью вида и величиной выживания.

Таблица 99. Коэффициент промыслового возврата у некоторых рыб (от отложенной икры)
Таким образом, мне представляется, что у всех рыб имеет место более или менее четко выраженная связь между численностью родительского стада, качеством его половых продуктов и численностью потомства. Эта связь, вытекающая из единства организмов и среды, обычно прямая, но в специфических условиях может оказаться и обратной.
Наличие прямой связи указывает, что регуляторные механизмы популяции работают нормально. При перегибе кривой действие регуляторных приспособлений нарушено, и они не в состоянии привести темп воспроизводства в соответствие с условиями жизни, в первую очередь с обеспеченностью популяции пищей. Обычно и при минимальных, и при максимальных плодовитостях стада отклонения от средних оказываются большими, чем при оптимальных. Наименьшая амплитуда изменчивости урожайности поколений и, следовательно, наибольшая устойчивость воспроизводства популяции наблюдается в верхнем отрезке восходящей части кривой, отражающей соотношение родители - потомство.
Пунктирные линии на рис. 95 указывают на то, что уровень выживания (определяемый соотношением напряженности связей в триотрофе) может быть различным и у одной популяции в разные периоды ее истории. Кроме того, у большинства популяций есть отдельные точки выживания, выходящие за пределы, свойственные для данной популяции. Обычно у популяций, живущих ближе к биологической границе ареала вида, таких точек оказывается больше. Эти точки отражают отклонения, вызванные появлением факторов или градиентов факторов, к воздействию которых популяция оказывается неприспособленной, или, наоборот, резким улучшением условий жизни. В первом случае это приводит к появлению точек, сильно отклоняющихся вниз от кривой, во втором - вверх от нее.
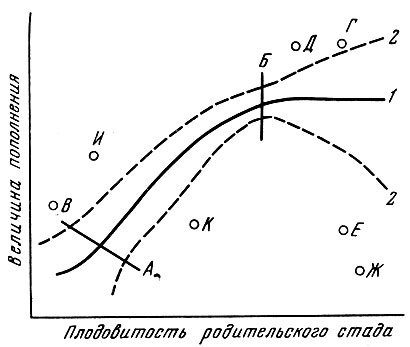
Рис. 95. Схема соотношения плодовитости родительского стада и численности потомства: 1 - средние показатели; 2 - возможные отклонения в пределах амплитуды приспособленности; А, Б - участок прямой связи; В, Г, Д, Е, Ж, И, К - отклонения, вызванные градиентами факторов к которым вид не приспособлен
Расхождение исследователей во мнениях о характере связей родителей и потомства, мне думается, зависит не только от ошибок в анализе фактического материала, но и от того, что большинство лиц, отрицающих наличие закономерной связи между численностью родителей и численностью потомства (Дементьева, Бойко, Земская, Танасийчук и др.), исследовало рыб (главным образом полупроходных), на размножении которых очень сильно сказалась деятельность человека, или рыб с очень сильно выраженными флюктуациями урожайности. Связи численности родительского стада и потомства нарушаются также у тех рыб (например, у атлантического лосося в Канаде), у которых численность молоди ограничивается определенным количеством убежищ, где она может прятаться от хищников, или численность которых (например, у многих камбал) ограничивается величинами кормовых площадей для рыб, переходящих к донному образу жизни. Таким образом, у большинства видов рыб, живущих в условиях ненарушенного воспроизводства, наблюдается более или менее четко выраженная связь численности родительского стада и потомства.
Мне думается, что на основе изложенного выше можно принять схему связи между численностью родительского стада и потомства (см. рис. 95). В определенных пределах обычно существует прямая корреляция между численностью родительского стада и численностью потомства, причем при минимальных численностях разброс отклонений от средней будет больше, чем при средних. При достижении родительским стадом определенной плодовитости (естественно, разной у различных видов, у одного вида в различных частях ареала и у одной популяции в разные годы), если при этом соответственно не изменятся обеспеченность пищей и условия жизни потомства, - прямая корреляция нарушается или даже переходит в обратную, причем опять как при минимальных, так и при максимальных численностях родительского стада размах уклонений оказывается много больше, чем при оптимальных, для этого плеча кривой.
Таким образом, закономерности, которым подчиняется связь производительности родительского стада и численности потомства, выражены достаточно четко и включаются в систему регуляторных механизмов, свойственных популяциям животных организмов.
Представление о наличии закономерной связи между производительностью родительского стада и численностью потомства вытекает из представления о приспособительности всех видовых свойств, т. е. отражает единство вида и среды.
Правильное понимание закономерностей, которым подчиняется взаимосвязь родительского стада и потомства, кроме большого теоретического интереса, имеет и важнейшее практическое значение. Оно важно как для правильной организации эксплуатации стад промысловых рыб, так и для обеспечения их воспроизводства. Разработку этого вопроса необходимо форсировать, обеспечив постановку глубоких исследований, базирующихся на правильных теоретических представлениях. Попытки решать этот вопрос на основе только поверхностных сопоставлений не могут дать нужных для нашего хозяйства результатов.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'