Смертность рыб от старости
Смерть от старости свойственна всем организмам. Если бы старые, потерявшие способность к размножению особи не погибали, а продолжали существовать, то жизнь вида очень скоро прекратилась бы. Смерть от старости в определенном возрасте - это видовое приспособительное свойство. В пределах популяции предельный возраст может несколько меняться в связи с изменением обеспеченности пищей. Как было показано выше, рыбы, созревшие в более молодом возрасте, обладают и меньшей продолжительностью жизни. Предельный возраст у рыб - это такое же видовое свойство, как и общий ход смертности особей в популяции. Естественно, ход смертности может несколько меняться в пределах определенной амплитуды, но общий ход кривой смертности остается специфичным для вида. Ниже мы подробнее остановимся на рассмотрении влияния вылова на ход "кривой смертности" и на структуру популяции.
Видовая специфичность характера кривой смертности показана на рис. 52. У одних видов рыб особи погибают тотчас после первого нереста, причем гибнут все самцы и самки (дальневосточные лососи рода Oncorhynchus, некоторые бычки Gobiidae). У ряда видов рыб погибают только все самцы, например у Callionymus lyra, или самцов гибнет значительно больший процент, чем самок, как это происходит у атлантического лосося. Таким образом, специфичен для вида не только общий характер смертности, но и характер смертности разных полов. Приспособительная сущность смерти старых особей заключается не только в том, что они освобождают "место в природе" последующим поколениям, но часто она имеет и другое значение. Так, меньшая продолжительность жизни самцов, чем самок, важна для повышения продуктивности популяции, так как популяционная плодовитость резко возрастает при увеличении массы самок (поэтому биологически целесообразен повторный нерест). Эффективность же осеменения икры мало зависит - естественно до определенных пределов - от размеров самцов. В то же время количество корма, которое потребовалось бы для обеспечения пищей повторно нерестующих самцов; оказывается весьма значительным, и его расход на поддержание жизни повторно нерестующих самцов сильно снизил бы общую продуктивность популяции.
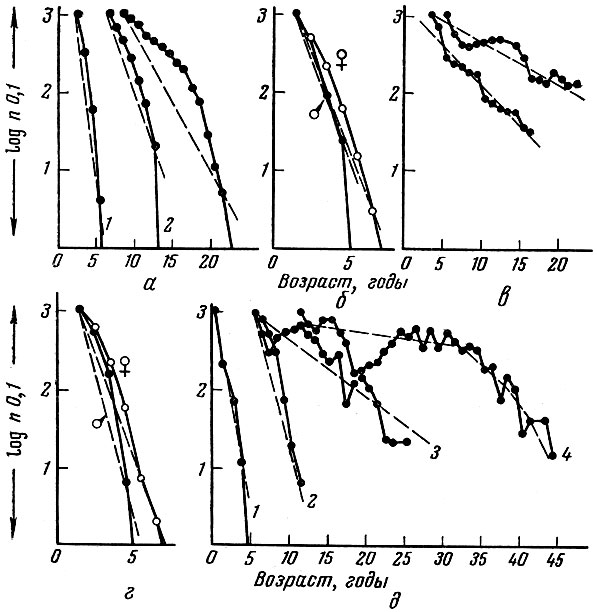
Рис. 52. Некоторые примеры кривых выживания в сравнительно малооблавливаемых популяциях (по Beverton a. Holt, 1959): а - сельдевые: 1 - Sprattus sprattus (L.), 2 - Clupea harengus pallasi Val., 3 - Clupea harengus harengus L., 6 - Cottus gobio L.; в - разные популяции обыкновенного окуня Perca fluviatilis (L.); г - Callionymus lyra; д - представители разных отрядов: 1 - Leurestes tenuis, 2 - Leucichthus kiyi, 3 - Coregonus clupeiformes, 4 - Acipenser fulvescens Raf
Наконец, смерть производителей в реках, особенно у лососей, нерестующих в малокормных речках предгорного типа, - это важнейший источник повышения кормности водоемов. О масштабе этого явления могут дать представление следующие цифры. Если общий улов лососевых в Амуре, например, составлял около 300 тыс. ц, а пропуск и изъятие относятся как 1 : 1, то это значит, что ежегодно в нерестовые речки бассейна Амура вносилось около 300 тыс. ц органических удобрений весьма высокого качества. Часть из этих пищевых ресурсов (вероятно, не более 10%) используется молодью в пищу непосредственно, а основная часть утилизируется кормовыми для молоди беспозвоночными. Принимая кормовой коэффициент трупов для молоди равным 10, можно установить, что трупами родителей непосредственно может прокормиться около 150 млн. шт. двухграммовых мальков. Если считать, что молодь лососевых является третьим звеном пищевой цепи (при кормовых коэффициентах на первом звене: труп - беспозвоночные 20 и беспозвоночные - молодь лососей 10), то, потребляя беспозвоночных, развивающихся на питании трупами, может прокормиться 67,5 млн. шт. молоди. Итого за счет трупов родителей может прокормиться в нерестовых реках 67,5 + 150 млн., т. е. около 217,5 млн. мальков. Даже если наши расчеты завышены, то все же видно, какое огромное количество пищевых ресурсов добавляют родительские особи из моря в нерестовые реки для молоди.
При усилении роли искусственного разведения в воспроизводстве стад дальневосточных лососей использованию "снёнки", т. е. трупов отнерестовавших особей, в качестве корма для выращиваемой молоди надо уделить больше внимания.
К сожалению, закономерности, которым подчиняются изменения предельного возраста рыб как в пределах одной популяции, так и у разных популяций одного вида и, наконец, у разных видов рыб изучены далеко не достаточно.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'