Причины смертности
Каждый вид характеризуется определенным максимальным предельным возрастом, которого могут достигать особи, его слагающие. Эта продолжительность жизни может меняться в известных пределах в связи с тем, в каких условиях живет популяция, и регулируется через физиологическое состояние особей условиями жизни.
Однако от старости умирает лишь очень небольшой процент особей у большинства видов, основная же часть популяции гибнет от других причин. Эта смертность, вызываемая самыми различными причинами, компенсируется плодовитостью особей. Чем выше плодовитость Особей вида, тем к более интенсивной смертности вид приспособлен. В известной степени условно все возможные причины смертности рыб можно подразделить:
1) от старости, включая посленерестовую смертность;
2) под воздействием абиотических условий;
3) от хищников, паразитов и болезней;
4) от нарушения обеспеченности пищей;
5) в результате вылова.
Эти причины взаимосвязаны, и такое подразделение в некоторой степени искусственно.
Обычно многие исследователи (Баранов, 1918, Beverton a. Holt, 1957; Тюрин, 1962 и мн. др.) подразделяют смертность на две большие категории: естественную (пп. 1, 2, 3 и 4) и промысловую смертность (п. 5). П. В. Тюрин (1962) так определяет эти понятия: "общая смертность" - это "суммарная убыль части популяции в результате вылова и естественной смертности". "Естественная смертность - гибель части популяции, вызываемая всеми другими причинами, кроме промысла". "Смертность от промысла - убыль части популяции в результате ее изъятия промыслом".
Под величиной общей смертности обычно понимают разность численности стада или той или иной его возрастной группы в начале и конце определенного отрезка времени. Соответственно величина естественной и промысловой смертности - это начальная численность стада минус количество погибших от естественных причин или выловленных рыб за определенный отрезок времени. Иногда естественная смертность подразделяется на зависящую от плотности популяции (density dependent) и независящую от плотности популяции (density independent). Однако последнее деление весьма условно. Система отношений вида и среды - это взаимосвязанная система, и взаимодействие вида с элементами абиотической среды оказывается различным в популяциях разной численности. Например, в годы мощных ходов дальневосточных лососей в нерестовые реки при одних и тех же зимних температурах и осадках относительно больше икры гибнет в результате промерзания нерестовых бугров, чем в годы, когда нерестовое стадо менее мощное. При одинаковых колебаниях уровня воды у большого нерестового стада гибнет от обсыхания больше икры, чем у небольшого нерестового стада, так как при незначительной численности нерестового стада производителями осваиваются более удобные нерестовые участки, чем в случае, когда в нересте участвует мощное стадо. Однако большее нерестовое стадо интенсивнее перекапывает грунт, увеличивая тем самым площадь нерестилищ и несколько улучшая условия инкубации икры.
Для каждого вида специфична не только общая величина смертности, но и ее распределение по отдельным возрастным группам и этапам развития. У одних видов наибольшая гибель приходится на стадию икринки, у других на этапы свободного эмбриона, у третьих на этап смешанного питания или более поздние этапы. Так, у дальневосточных лососей основная смертность падает на период жизни в буграх на стадии икринки и свободного эмбриона. У многих стад атлантического лосося и форелей наибольшая гибель приходится на первое лето жизни в реке после выхода из нерестовых бугров: у сельдей, анчоусов, трески и многих других рыб - на этап смешанного питания и, наконец, у многих амурских пелагофильных рыб наибольшая смертность приходится на момент перехода от жизни в толще воды русла реки к жизни в прибрежной зоне.
Различными оказываются и причины, вызывающие массовую гибель рыб на разных этапах онтогенеза. На стадии икринки и свободного эмбриона ведущие отношения и основные причины, определяющие гибель,- это абиотические условия, в первую очередь условия дыхания, а также воздействие хищников (Крыжановский, 1948, 1949). С переходом на внешнее питание и приобретением личинкой способности активного движения летальное действие абиотических условий обычно снижается; на первое место выступает влияние обеспеченности пищей и большое значение как фактор смертности сохраняют хищники. При этом на первых этапах внешнего питания недостаток пищи часто непосредственно может приводить к гибели особей, а на более поздних этапах его действие обычно сказывается косвенно - через затормаживание роста, удлинение периода активности в связи с необходимостью тратить больше времени на поиск пищи, вследствие чего рыба становится более доступной для хищников. Наконец, в связи с деятельностью человека включается такой мощный фактор смертности, как вылов. У разных видов рыб вылов приходится на разные этапы онтогенеза, но обычно основное воздействие вылова приходится на период жизни рыбы после достижения половой зрелости.
При загрязнении человеком мест естественного размножения рыб и иных отклонений от нормальных условий, а также несовершенстве биотехники рыбоводства иногда довольно значительная часть икры, свободных эмбрионов и личинок гибнет в результате появления у них дефектов в строении и нарушений процесса развития.
Причины, определяющие процесс динамики популяции, и ведущие связи популяции со средой можно схематически изобразить следующим образом (рис. 51), Как видно на схеме и как об этом говорилось выше, количество и жизнестойкость пополнения нерестовой популяции в значительной степени определяются количеством и качеством половых продуктов, производимых родительским стадом. Это зависит как от условий нагула родительского стада, так во многих случаях и от соотношения впервые нерестующих, повторно нерестующих и старых рыб, качество производимого потомства у которых далеко не одинаково. На стадиях икринки и свободного эмбриона ведущее значение имеют абиотические влияния и хищники; на этапе смешанного питания включается такой мощный фактор, как обеспеченность пищей, который сохраняет свое значение на всех последующих этапах онтогенеза.
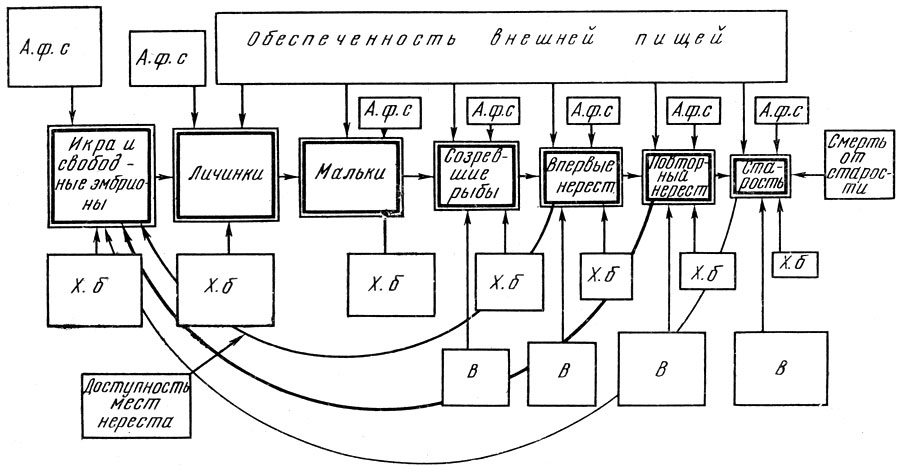
Рис. 51. Схема основных причин, определяющих процесс динамики популяции, и ведущие связи популяции со средой. Относительная величина прямоугольников условно характеризует интенсивность воздействия того или иного фактора или относительную численность популяции. Толщина стрелок, идущих от половозрелой части стада к икре, характеризует относительную роль в воспроизводстве впервые, повторно нерестующих и старых рыб: А. ф. с - абиотические факторы смертности; X. б - хищники и болезни; В - вылов
Роль хищников на последующих этапах становится все меньше, хотя может быть тоже весьма значительной. У промысловых рыб на смену влияния хищников включается со все нарастающей силой такой фактор, как вылов.
Я здесь специально не выделяю косвенных форм воздействия человека, очень существенно изменяющих условия жизни рыб и часто сильно влияющих на величину смертности. Однако все косвенные формы воздействия человека сказываются или через изменение абиотических условий, или через изменения обеспеченности пищей, или через изменения действия хищников, паразитов и болезней.
Методика определения смертности, к сожалению, еще очень слабо разработана.
Для учета смертности на ранних этапах онтогенеза, обычно, проводятся периодические просчеты икры, свободных эмбрионов и личинок в единице объема воды или на единицу площади дна. По разнице в последующих ловах определяется интенсивность убыли.
Прямые определения величины общей смертности осуществимы в довольно редких случаях, когда имеется возможность полного облова водоема из года в год и учета всех изменений, происходящих в популяции. Подобного рода исследование выполнено, например, Альмом (Aim, 1952) на двух популяциях окуня.
В настоящее время для оценки общей смертности обычно применяются две группы методов: 1) анализ возрастного состава популяции и 2) массовое мечение и учет возврата меток. К сожалению, обе группы методов пока не позволяют точно определить величину общей смертности и дают лишь грубо приблизительные цифры. Однако пока что во многих случаях они являются единственными.
Наиболее точный способ заключается в сопоставлении численности поколения определенного года в уловах неселективными орудиями лова за ряд лет, равных продолжительности жизни поколения. Принимая средний улов поколения на единицу рыболовного усилия за 100% и вычитая из него улов этого поколения за следующий год, выраженный в процентах улова предыдущего года, мы получаем смертность за год.
Если иметь наблюдения за ряд лет, равный продолжительности жизни рыбы, можно получить картину общей смертности. При относительной стабильности интенсивности рыболовства определить общую смертность поколения можно и по общим уловам данной облавливаемой популяции* Однако практически пользоваться этим методом довольно сложно в отношении многих рыб, а в отношении некоторых вообще невозможно и требует в случае рыб с длинным жизненным циклом продолжительного срока. Поэтому во многих случаях для определения общей смертности исследователи пользуются анализом возрастного состава, полагая, что правое "плечо" кривой возрастного состава улова отцеживающими орудиями лова отражает для старших возрастных групп соотношение возрастных групп в популяции. При этом делается допущение, что начальная величина поколений из года в год одинакова.
| Возвраст, годы | Коэффициент убыли, % |
|---|---|
| 7 | 12 |
| 8 | 23 |
| 9 | 33 |
| 10 | 32 |
| 11 | 34 |
| 12 | 39 |
| 13 | 27 |
| 14 | 45 |
Однако известно, что численность поколений у рыб, особенно тех, у которых выражены флюктуации, может меняться в десятки раз; следовательно, и ошибки в определении общей смертности могут также выражаться цифрами порядка десятков раз. У рыб, у которых флюктуации выражены слабо, соответственные ошибки будут меньше. На основании анализа многолетних данных о возрастном составе популяций современного азовского судака и судака из кухонных остатков древнего человека Е. Г. Бойко (1964а) установил следующую интенсивность естественной смертности не промышляемой популяции: Однако переносить эти данные на современного судака надо с большой осторожностью. Предельный возраст рыб в кухонных остатках древнего человека мог быть иногда выше современного, что связано с более медленным ростом и более поздним наступлением половой зрелости. Эти цифры можно принять для первой ориентировки, но едва ли можно согласиться с Е. Г. Бойко, что с повышением интенсивности рыболовства эти потери могут не приниматься в расчет. Те потери от естественной смертности, которые приводит Е. Г. Бойко, - это, конечно, не только смерть от старости, но и гибель от многих других причин. Конечно, по мере продвижения к предельному возрасту, процент рыб, умерших от старости, будет нарастать, но, вероятно, даже у рыб предельного возраста он будет менее 100%.
Определяя общую смертность по среднему возрастному составу стада, основанному на многолетних данных, мы получаем и средние показатели смертности. В то же время даже по тем незначительным надежным данным, которые имеются, общая смертность так же, как и величина пополнения, может весьма сильно меняться из года в год, причем в отношении многих видов рыб это справедливо и не только для первого года жизни. Поэтому определение общей смертности по возрастному составу стада возможно лишь по средним многолетним данным и лишь для первой ориентировки главным образом в отношении рыб со слабо выраженными флюктуациями. Более подробно методика оценки общей смертности рассмотрена П. В. Тюриным (1962, 1963, 1967, 1972).
В работах П. В. Тюрина, представляющих большой теоретический и практический интерес, как мне представляется, недоучитывается связь величины предельного возраста, достигаемого особью, с возрастом достижения ею половой зрелости и темпом ее роста (см. выше). Мне думается также, что едва ли целесообразно вычисление общего коэффициента смертности. Коэффициент смертности, понимаемый как процент погибающих особей за данный отрезок времени по отношению ко всем особям популяции, реален для каждой возрастной группы.
Средний же коэффициент, по-моему, не характеризует процесса убыли. Он только показывает, насколько велика продолжительность жизни рыб данного вида, что известно и без него, хода же смертности он не характеризует.
Некоторые авторы, исходя из представления, что относительная смертность пропорциональна численности стада, вводят так называемый коэффициент мгновенной смертности - Z, выражаемый формулой
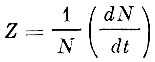 (7)
(7)где:
N - численность стада;
t - время (см. Засосов, 1970).
Однако мне представляется, что этот показатель не отражает реального биологического явления. Тем более что и исходная посылка в большинстве случаев является неправильной.
В некоторых случаях величину общей смертности можно определять по возврату меченых рыб. Принцип определения смертности методом мечения заключается в следующем: принимается, что величина смертности в популяции за определенный отрезок времени соответствует снижению в уловах числа помеченных рыб за этот промежуток (Parrish, no Graham, 1956; Ricker, 1958). Однако, учитывая погрешности техники мечения и изменение поведения помеченных особей промысловых рыб, а главное разную смертность рыб с метками и без них (Parker, Black a. Larkin, 1963), пользоваться методом мечения при оценке как общей, так и. промысловой смертности можно лишь как подсобным приемом, который контролировался бы другими методами.
Очень важный вопрос - это установление относительной величины разных форм смертности, в первую очередь отчленение естественной смертности от промысловой. К сожалению, до настоящего времени этот вопрос, несмотря на то что ему посвящена довольно обширная литература, все еще остается нерешенным. Лишь в некоторых случаях удается установить относительные величины отдельных форм смертности, как это, например, удалось сделать К. Р. Фортунатовой (1961) в отношении воблы Северного Каспия. К. Р. Фортунатова показала, что в 1953 г. хищниками в Северном Каспии было съедено воблы лишь немногим меньше (447 тыс. ц), чем добыто промыслом (580 тыс. ц). За 1948-1953 гг. процент воблы, съедаемой хищниками, колебался по отношению к вылову от 72 до 93%.
В настоящее время существуют три основных метода определения относительной величины естественной и промысловой смертности. Первый основывается на сопоставлении величины промысловой смертности, определенной по возврату меченых рыб и изменений численности, установленной по динамике возрастного состава. Второй метод основывается на сопоставлении величины общей смертности и интенсивности рыболовства. Наконец, третий основан на приблизительном определении величины естественной смертности и сопоставлении ее с общей смертностью (Cushing, 1968).
К сожалению, общего надежного метода определения величины разных форм смертности мы пока не имеем. Сопоставление кривых возрастного состава популяции при разных интенсивностях промысла (Beverton a. Gulland, 1958) или сравнение возрастного состава не облавливаемых и облавливаемых популяций (Бойко, 1964а), по моему убеждению, не дает достаточно надежных оснований для выделения промысловой смертности из естественной. Тем более сложно определение величины естественной смертности и ее разных форм.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'