Типы нерестовых популяций у рыб и их динамика
Как видно из изложенного выше, по возрастной структуре популяции виды рыб весьма разнообразны. Возрастная структура иногда существенно различается и у разных группировок в пределах вида. Стадо каждого вида рыб слагается из особей, различных по физиологическому состоянию, в частности по половой зрелости. У большинства рыб популяция состоит из: неполовозрелых особей; рыб, впервые достигших половой зрелости; повторно нерестующих, и рыб старых, частично или полностью утративших способность воспроизводства. В пределах каждой из этих групп рыбы в свою очередь находятся на разных этапах развития. Однако соотношение этих Отдельных групп особей в популяции, структура каждой из этих групп весьма различны, специфичны в определенных пределах для каждого вида и в то же время могут изменяться в связи с изменением условий жизни.
Г. Н. Монастырский (1949, 1953), анализируя структуру нерестовой части популяции, выделяет три "типа нерестовых популяций".
Первый тип - это популяция, у которой отсутствуют повторно нерестующие и старые, потерявшие способность размножения, особи. Все половозрелые рыбы после первого размножения погибают. К этому типу относятся дальневосточные лососи, некоторые бычки. В пределах этого типа существенные различия наблюдаются не только в размерах, при которых рыбы достигают половой зрелости, но и в возрастной структуре впервые созревающих рыб. Бычки (Aphya, Bethophilus) достигают половой зрелости все в одном возрасте, а поколения дальневосточных лососей созревают в течение нескольких лет. Так, у кеты обычно нерестовое стадо состоит из рыб в возрасте от 2+ до 7 + , у красной от 3+ до 6+, у горбуши от 1+ до 2+ и т. д. Такая разница в структуре половозрелой части популяции связана с различной стабильностью условий размножения. Стадо горбуши почти полностью исчезнет, если погибнут два поколения. Для кеты и других дальневосточных лососей необходимо значительно больше неурожайных лет, чтобы стадо перестало существовать. Как было отмечено мною (Никольский, 1952), условия нереста горбуши действительно значительно стабильнее условий нереста кеты. В то же время рыбы, у которых половая зрелость наступает в более раннем возрасте, могут компенсировать большую гибель половозрелой части стада. Так, по данным Нива (Neave, 1953), стадо горбуши может выдерживать изъятие более 60% половозрелых особей без сокращения численности потомства, а стадо кеты - около 50%.
Второй тип нерестовой популяции состоит как из пополнения, т. е. впервые нерестующих рыб, так и рыб повторно нерестующих, а иногда и рыб старых, потерявших способность к размножению. Как указывает Г. Н. Монастырский (1953), ссылаясь на А. А. Махмудбекова (1947), в стаде каспийского пузанка попадаются старые особи размером до 34 см в возрасте 10 лет, потерявшие способность воспроизводства. Характерной особенностью этого типа нерестовой популяции является, по Монастырскому, то, что в нерестовом стаде относительная величина пополнения больше остатка.
К рыбам этого типа нерестовой популяции относятся каспийские сельди, атлантический лосось и ряд других видов рыб. И в пределах этого типа у разных видов также есть существенные различия в структуре стада и времени полового созревания.
Третий тип нерестовой популяции характеризуется преобладанием остатка над пополнением. У рыб этой группы, как правило, время полового созревания растянуто, структура остатка часто весьма сложна. Остаток состоит как из рыб, нерестующих каждый год, так и из рыб, пропускающих один или несколько нерестовых сезонов (например, многие осетровые). У рыб с популяцией этого типа часто довольно большой процент составляют старые рыбы.
Однако еще В. Н. Майский (1953) на примере тюльки и хамсы показал, что у одного и того же вида рыбы в разные годы "тип нерестовой популяции" может быть различным: в одни годы пополнение оказывается больше остатка, в другие наоборот.
Так, у хамсы Черного моря впервые нерестующие рыбы составляли в разные годы (с 1952 по 1962) от 26 до 97% от нерестового стада (Павловская, 1963). Подобная же картина наблюдается у воблы (относимой Монастырским к рыбам с популяцией третьего типа) (табл. 72).

Таблица 72. Соотношение пополнения и остатка в стаде воблы северного каспия за разные годы (по Монастырскому, 1949)
Таким образом, в 1934 и 1935 гг. вобла имела нерестовую популяцию третьего типа, а в 1936 и 1937 гг. - второго. Еще более резкие различия по годам наблюдаются у рыб с более значительными, чем у воблы, колебаниями урожайности поколений.
Ю. Е. Лапин (1961, 1971) и Л. С. Иванов (1963) отмечают, что структура популяции весьма динамична; она определяется соотношением пополнения и остатка, которое непрерывно меняется, отражая самую динамику численности стада.
Г. Н. Монастырский (1949) правильно указал на видовую специфичность структуры нерестовых популяций у рыб, но неверно принял стабильность границ между выделенными им типами. Несомненно, что границы между намеченными Г. Н. Монастырским типами нерестовых популяций условны не только между популяциями второго и третьего, но и первого и второго типов. Так, например, у атлантического лосося известны случаи 100%-ной гибели производителей после нереста.
Кроме того, надо иметь в виду, что биологическая специфика популяции едва ли изменится, будет ли остаток равен 1 или 0%. Из математических моделей, приводимых Ю. Е. Лапиным (1961, 1971), видно, что изменение величины пополнения в 5 раз может изменить соотношение пополнения и остатка от 1 : 2 к 5 : 2, а выше мы видели, что относительная численность поколения может меняться у некоторых видов во многие десятки раз. Однако, хотя границы выделенных Г. Н. Монастырским типов нерестовых популяций и условны, несомненно, что структура популяции и характер ее динамики отражают специфику взаимосвязей популяции со средой. Каждая популяция слагается из ряда поколений, которые рождаются, растут, созревают, воспроизводят один или несколько раз в жизни себе подобных, стареют и исчезают. Отдельные поколения в популяции воздействуют друг на друга, влияя на рост особей, ход созревания и смертность. Знать судьбу поколений, слагающих популяцию, необходимо как для понимания структуры популяции и ее изменений, так и для понимания динамики численности в целом.
В. Томпсон (Thompson, 1959) схематически изобразил кривую выживания и созревания одного поколения у трех биологических групп рыб, различающихся по продолжительности жизни и характеру полового созревания (рис. 40). Однако в схеме Томпсона выпали такие стороны жизни поколения, как соотношение впервые и повторно нерестующих и старых рыб.
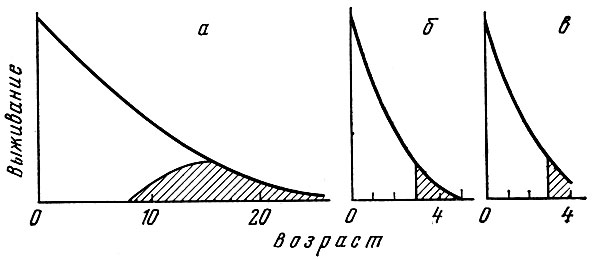
Рис. 40. Кривые выживания длинноцикловых (а), короткоцикловых (б) и лососевых (в) рыб (по Thompson, 1959). Штриховкой обозначен период половой зрелости
На приводимых схемах (рис. 41) "судьбы поколения", составленных в отношении рыбы с разным жизненным циклом, видно, что в первый период жизни все поколение состоит из неполовозрелых рыб; в следующий период количество неполовозрелых рыб убывает и постепенно или быстро сходит на нет. Поколение далее состоит только из зрелых рыб, к которым к концу жизни поколения все в большем количестве у рыб с повторным нерестом примешиваются и старые особи. Одновременно с ростом особей, созреванием поколения и его старением идет и его убыль. Ход кривых на схемах Томпсона отражает этот процесс. Однако судьбы поколений в пределах даже одной и той же популяции могут быть весьма различными. Во взаимодействии с внешними условиями регуляторные механизмы, действующие у популяций рыб, изменяют не только величину пополнения, но и соотношение отдельных групп в пределах половозрелой части стада. В некоторых крайних случаях изображенные на рис. 41 второй и третий случаи "жизни поколения" могут быть свойственны разным поколениям одной и той же популяции.
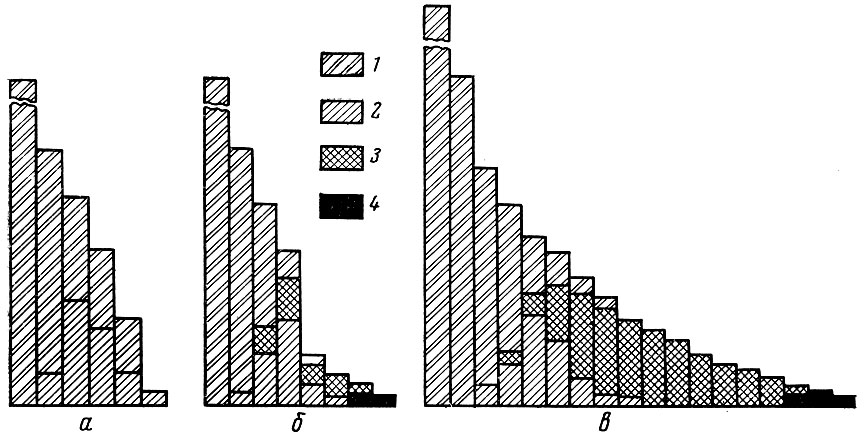
Рис. 41. Судьбы поколений у рыб с разной структурой стада. Динамика численности и созревания поколения рыб: а - без повторного нереста; б - рыб с повторным нерестом при высокой обеспеченности пищей; в - рыб с повторным нерестом при низкой обеспеченности пищей; 1 - неполовозрелые рыбы; 2 - созревшие; 3 - повторно нерестующие; 4 - старые
Схема вторая отражает случай, когда поколение живет при высокой обеспеченности пищей, рано созревает и раньше достигает старости.
Третья схема характеризует поколение, находящееся в менее благоприятных условиях обеспеченности пищей, позднее созревающее, но обладающее большей продолжительностью жизни.
Если численность поколения с момента откладки икры до гибели последней особи непрерывно убывает, то биомасса поколения, как правило, изменяется несколько иначе и максимальная биомасса приходится на более старшие возрасты, различные у разных видов рыб. Попытаемся на примере салаки, корюшки и судака проследить динамику численности и биомассы отдельных поколений.
У салаки Clupea harengus membras (L.) Финского залива (рис. 42) наибольшая биомасса приходится на вторую возрастную группу (1+).
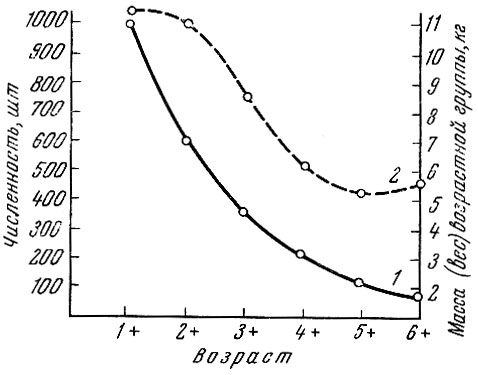
Рис. 42. Численность (1) и биомасса (2) салаки Финского залива в последовательные годы жизни (по Тюрину, 1963)
График динамики численности и биомассы одного поколения корюшки Кандалакшского залива (поколение 1958 г.) показывает, что если численность из года в год падает (смертность от третьего к четвертому году 38%, от четвертого к пятому году 98%), то биомасса популяции от третьего к четвертому году увеличивается почти на 25% и только потом резко снижается (рис. 43).
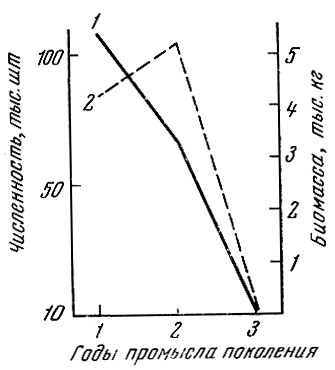
Рис. 43. Численность (1) и биомасса (2) корюшки Кандалакшского залива Белого моря поколения 1958 г. рождения в последовательные годы промысла (по материалам Т. Н. Беляниной)
У судака Lucioperca lucioperca (L.) оз. Ильмень (Тюрин, 1963) биомасса и численность меняются в разные годы в связи с неодинаковой плотностью популяции и темпом роста (рис. 44). В 1936 г., когда промысел был весьма интенсивным и судак рос быстро, максимальная биомасса возрастной группы приходилась на рыб в возрасте 7 + .
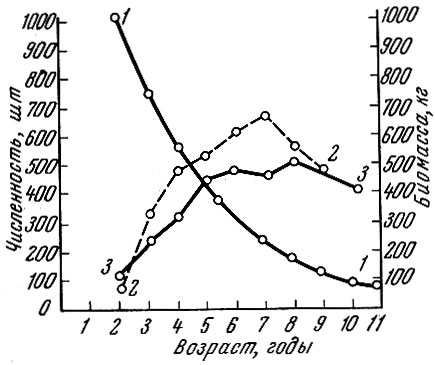
Рис. 44. Численность и биомасса судака оз. Ильмень в последовательные годы жизни (по Тюрину, 1963): 1 - численность; 2 - биомасса за 1936 г.; 3 - биомасса за 1946 г
В 1946 г. после частичного запуска в годы войны темп роста рыб сильно замедлился и максимальная биомасса переместилась на возрастную группу 8+.
Мы рассмотрели три типа весовой структуры популяции: 1) салаки Финского залива, у которой максимальная биомасса приходится на вторую возрастную группу; 2) корюшки, у которой максимальная биомасса приходится на третий год жизни, и 3) судака, у которого максимальная биомасса приходится на рыб в возрасте 7+, 8+. Таким образом, весовая структура популяции, обладая видовой специфичностью, меняется в известных пределах в связи с изменением условий жизни. Структура популяции - это как бы наложенные одно на другое поколения, число которых равно предельному числу лет, достигаемому рыбой. Динамика численности, роста, созревания и старения особей в поколениях и определяет структуру популяции. Структура популяции - это, как мы говорили, видовое приспособительное свойство, отражающее характер взаимодействия популяции и ее среды.
На изменения условий жизни популяция приспособительно реагирует изменением своей структуры и воспроизводительной способности. При улучшении обеспеченности пищей темп воспроизводства популяции возрастает, при ухудшении замедляется
Прежде чем перейти к рассмотрению приспособительного механизма саморегуляции структуры популяции, необходимо еще раз рассмотреть, приспособлением к чему она является.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'