Изменения возрастного состава в пределах одной популяции
Существенные изменения происходят в структуре популяций и в пределах одного и того же стада. Возрастной состав стада есть результат взаимодействия трех процессов: пополнения (урожайности), роста и убыли. От соотношения этих трех взаимосвязанных процессов и зависят изменения возрастной структуры как популяции в целом, так и ее половозрелой части. Большое влияние на возрастную структуру стада оказывает разница в численности (урожайности) отдельных поколений. У разных видов рыб степень урожайности разных поколений колеблется в разных пределах (рис. 28 и 29).
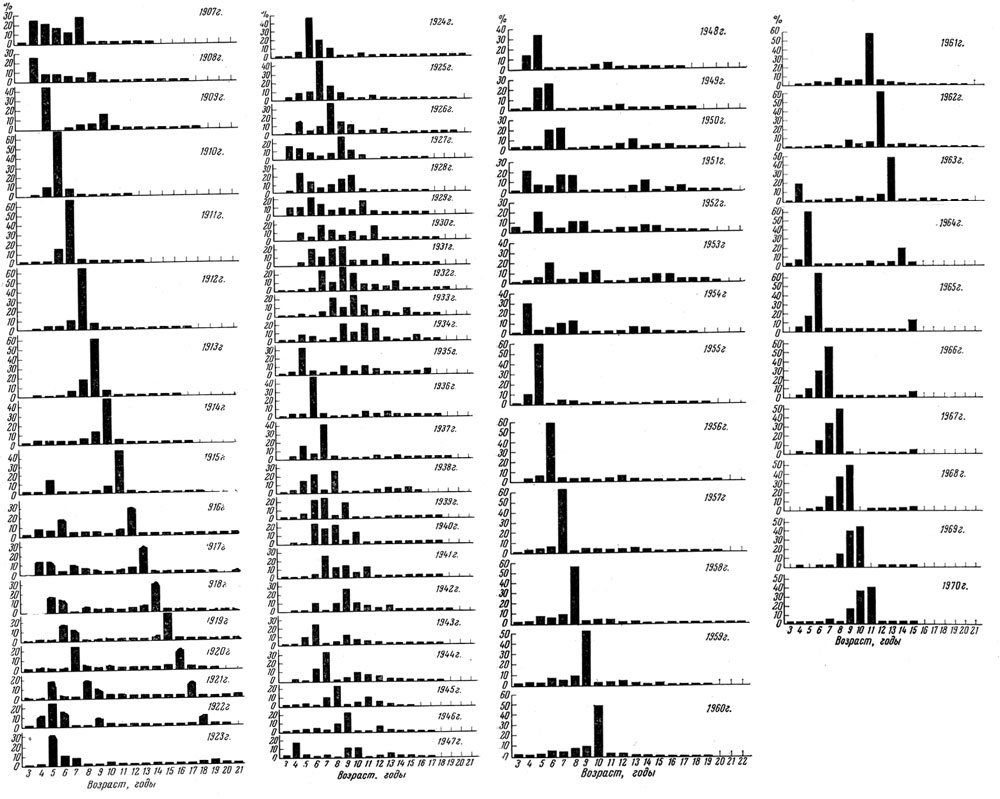
Рис. 28. Возрастной состав нерестового стада атлантическо-скандинавских сельдей
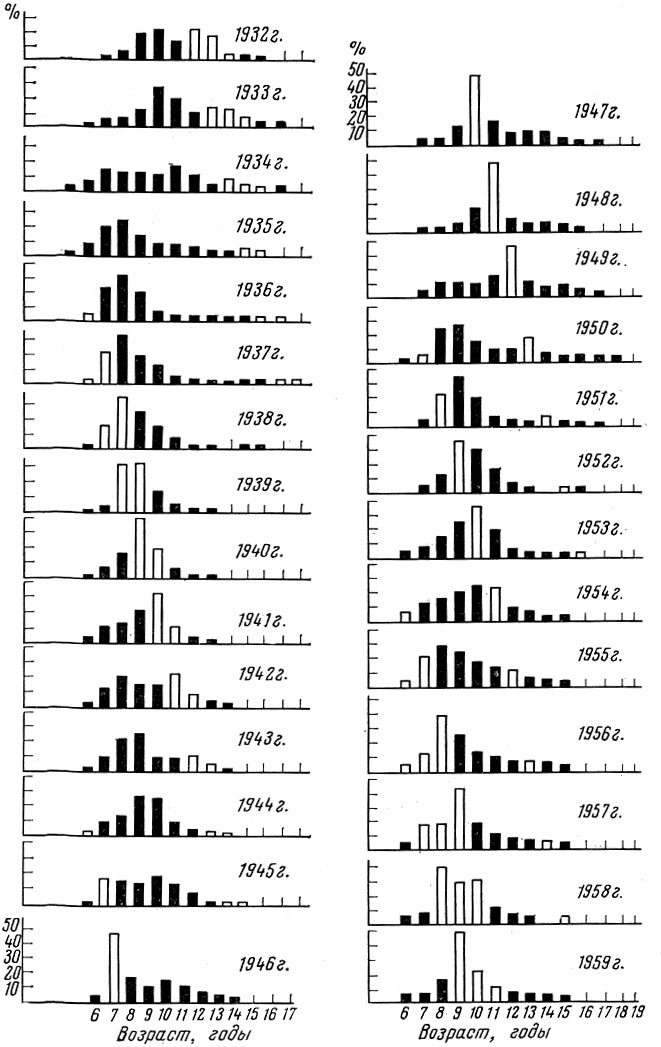
Рис. 29. Возрастной состав лофотенской трески (по Роллефсену, 1954; Марти, 1961)
У одних видов (треска, сельдь и др.) флюктуации, т. е. колебания численности отдельных поколений, выражены сильно, и в некоторых случаях мощное поколение может оказаться больше слабого в 50-90 раз. Такие колебания урожайности в первую очередь и определяют ту изменчивость возрастного состава половозрелой части популяции, какую мы наблюдаем, например, у аркто-норвежской трески и атланти-ческо-скандинавских сельдей (табл. 59). Конечно, урожайные поколения несколько рассредоточиваются при вступлении в нерестовое стадо вследствие разновременности созревания особей, но все же их влияние на возрастную структуру стада очень велико.
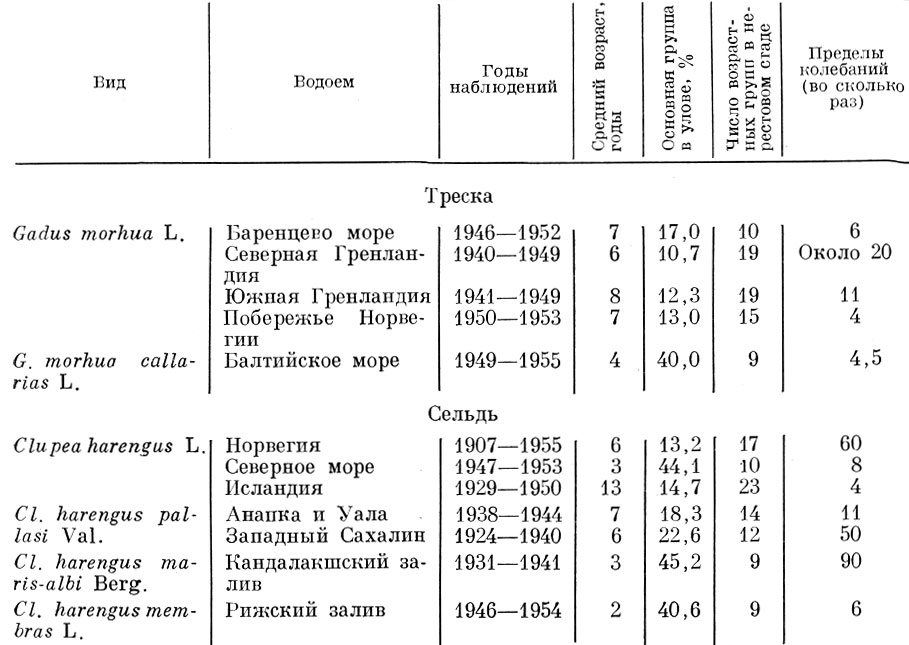
Таблица 59. Изменения основной возрастной группы в нерестовом стаде сельди и трески (по данным Annales biologiques и изданий Полярного и Тихоокеанского институтов рыбного хозяйства и океанографии)
У ряда видов рыб колебания урожайности по годам невелики. Вне воздействия рыболовства, т. е. изменения интенсивности смертности, возрастная структура стада у них почти не меняется (естественно, при условии стабильной обеспеченности пищей) (табл. 60).
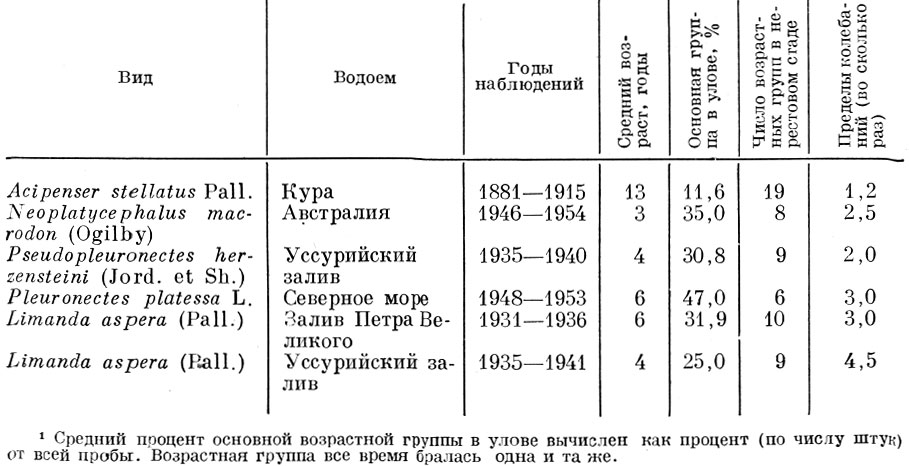
Таблица 60. Изменения относительной величины основной возрастной группы некоторых видов рыбsup1/sup (по данным Державина, 1922, 1952 Houston, 1955 и Моисеева, 1953)
Кроме колебаний урожайности, существеннейшее влияние на структуру популяций оказывает изменение обеспеченности пищей и роста рыб в популяции. В случае повышения обеспеченности популяции пищей, которое может быть вызвано увеличением кормовой базы, удлинением сезона нагула или разреживанием популяции, у рыбы ускоряется рост, раньше наступает половая зрелость, иногда меняется соотношение полов и обычно сокращается продолжительность жизни. Так, например, возрастной состав морской камбалы под влиянием интенсификации рыболовства и его снижения в годы первой и второй мировых войн менялся следующим образом: в годы интенсификации рыболовства ускорялся темп роста рыб и происходило омоложение стада; в годы военных запусков рост замедлялся и увеличивались как средний, так и максимальный возраст рыб в нерестовом стаде (рис. 30).
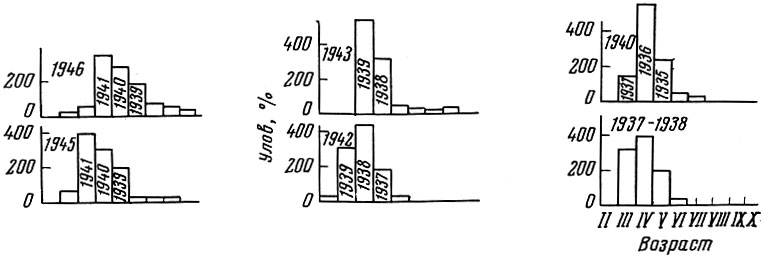
Рис. 30. Возрастной состав морской камбалы в промысловых уловах на Доггер-банке (по Jensen, 1948). Размеры рыб более 27 см; внутри столбиков указан год рождения поколения
Проникновение моллюска Dreissena в Учинское водохранилище улучшило условия питания плотвы, значительно ускорило ее рост и вызвало перестройку структуры популяции этого вида. По-видимому, у всех видов рыб существует определенная связь между возрастом и размерами полового созревания, а также предельным возрастом и размерами, которых рыба может достигать.
П. А. Дрягин (1934) показал, что у большинства рыб размеры, достигаемые особью ко времени полового созревания примерно вдвое меньше максимальных размеров, достигаемых рыбой. Следовательно, если рыба растет быстрее, она скорее достигает возраста половой зрелости и в более молодом возрасте достигает и предельных размеров; при этом, естественно, сократится и продолжительность жизни. Наоборот, замедление роста связано с более поздним достижением половой зрелости и большей продолжительностью жизни. Наконец, на возрастную структуру популяции существенное влияние оказывают характер и интенсивность смертности.
Так же, как возрастной состав, размерный состав и весовая структура популяции специфичны для вида и изменяются соответственно изменениям условий жизни.
У тех видов рыб, у которых разница в урожайности отдельных поколений значительна, очень сильно меняется по годам и размерный состав, и структура биомассы популяции. (Под структурой биомассы популяции я понимаю распределение биомассы по размерным или возрастным группам). Как это хорошо показано многими исследователями, у сельдей Северной Атлантики и северной части Тихого океана размерный состав только нерестовой популяции из года в год изменяется очень сильно. Это особенно отчетливо видно, если проводить анализ, пользуясь методом Оскара Зунда (Sund) или профилями Зунда, как их называет А. В. Морозов (1937). Как видно на рис. 31-34, размерный состав сельди и трески, даже если анализировать материал за относительно короткие отрезки времени, изменяется очень сильно. Меняется также соответственно и весовая структура популяции.
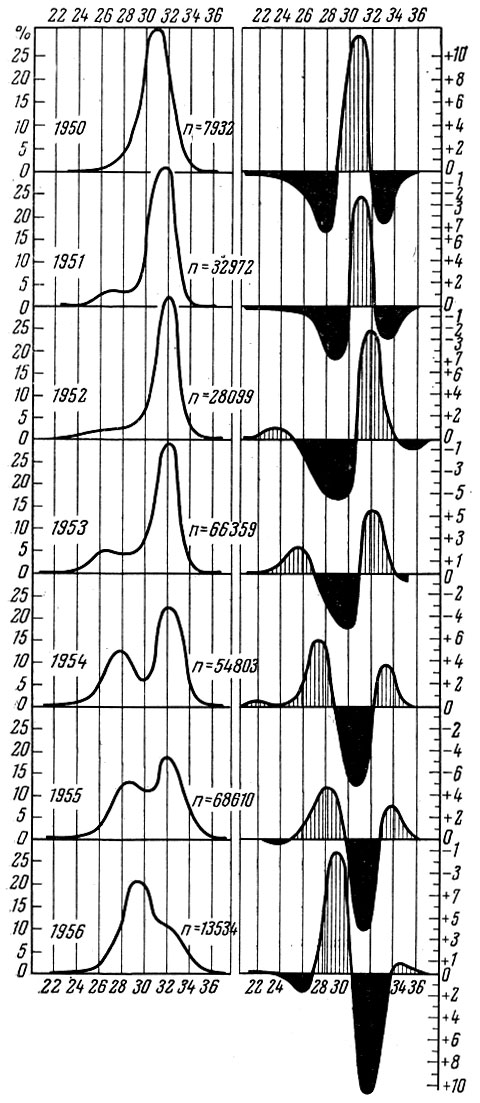
Рис. 31. Размерный состав атлантическо-скандинавских сельдей за 1950-1955 гг. (по Marty, 1958): слева - пределы колебаний; справа - отклонения от общей средней амплитуды
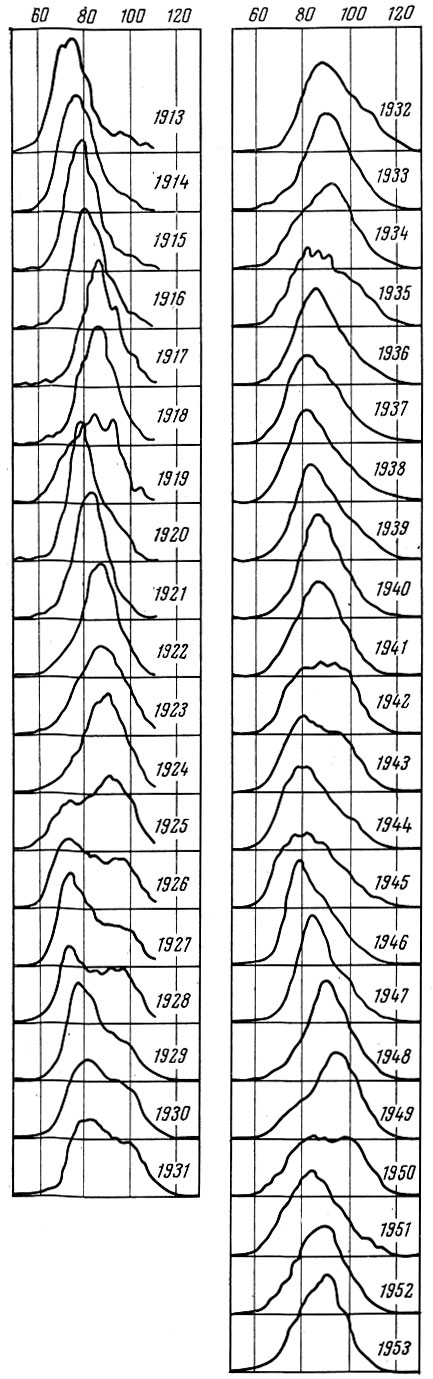
Рис. 32. Размерный состав лофотенской трески (по Rollefsen, 1954)
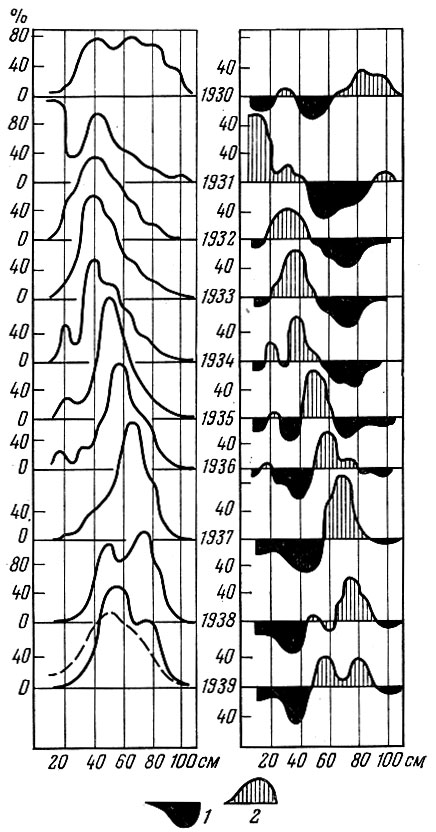
Рис. 33. Размеры трески в южной части Баренцева моря в 1930-1939 гг. (по Маслову, 1957): слева - кривые размеров трески за каждый год; справа - отклонения рядов за каждый год от среднего ряда размеров за 1930-1939 гг.; 1 - данные размеры предмтавлены в недостатке; 2 - в избытке по сравнению со средним многолетним рядом
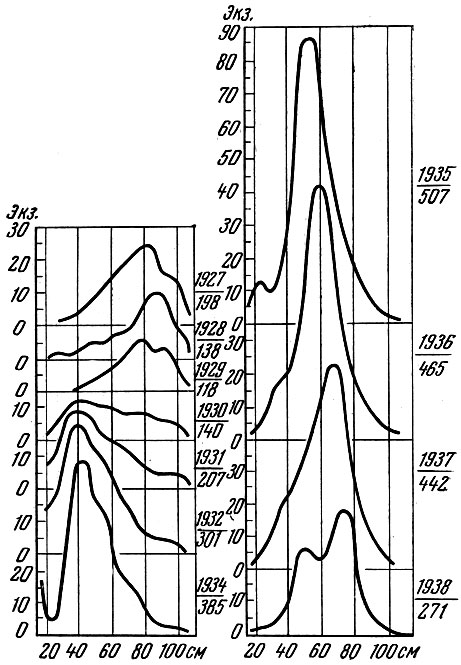
Рис. 34. Взвешенные ряды размеров трески южной части Баренцева моря за 1927-1938 гг. (по Маслову, 1957): в числителе - год наблюдений; в знаменателе - число рыб в среднем годовом улове за час траления
С другой стороны, размерная и весовая структура популяций, живущих в стабильных условиях обеспеченности пищей и урожайности поколений, остается почти неизменной из года в год. Примером может служить размерная и весовая структура популяции обыкновенного карася Carras-sius carassius (L.) в одном из подмосковных прудов, которая полностью облавливалась в одни и те же числа одиннадцать лет подряд (после анализа выпускалась обратно). Как видно из приводимых кривых (рис. 35 и 36), пока ее численность оставалась высокой (до 1966 г.), размерная и весовая структура ее не изменялась. При падении численности ниже определенного предела (в результате изъятия части особей) рост особей в популяции ускорился и ее размерная и весовая структура изменились.
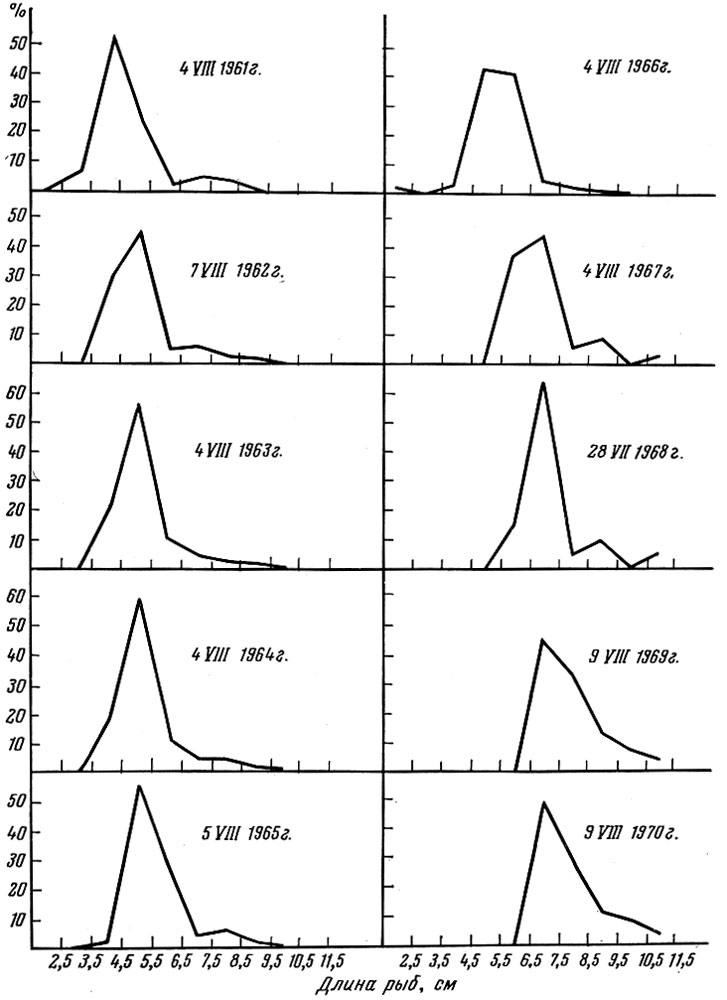
Рис. 35. Размерный состав (в %) популяции обыкновенного карася в 1960-1970 гг.(Указаны даты облова по Никольскому и Шубниковой, 1974)
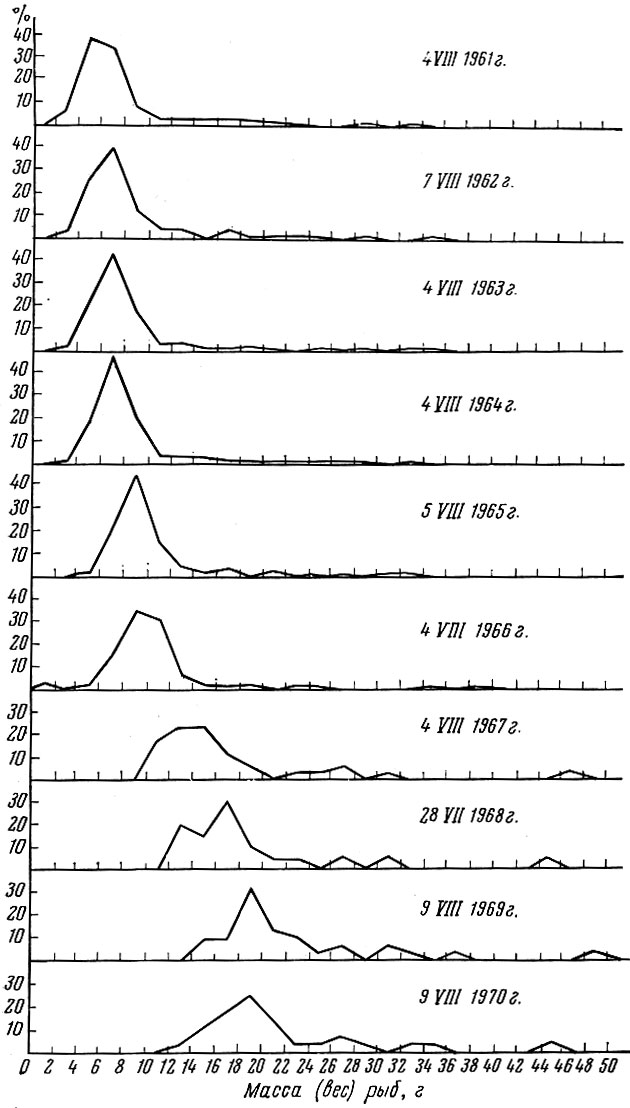
Рис. 36. Биомасса (в %) популяции обыкновенного карася в 1961- 1970 гг. (Указаны даты облова по Никольскому и Шубниковой, 1974)
Изменения размерного и весового состава популяции отражают изменения условий жизни. Расширение размерного ряда позволяет популяции осваивать более широкий спектр кормов и обеспечивать более стабильное пополнение. Сокращение размерного ряда возможно в условиях высокой обеспеченности пищей и связано с повышением интенсивности воспроизводства при относительно стабильных условиях размножения.
В пределах стада рыбы, особенно в высоких и умеренных широтах, обычно возникают временные, более или менее длительные, но редко существующие в течение всей жизни поколения, группировки - "элементарные популяции" по терминологии Н. В. Лебедева (1967). Эти группировки объединяют главным образом особей одного поколения, характеризующихся сходным биологическим состоянием (жирностью, зрелостью гонад и т. д.), размерным составом и поведением. Иногда в элементарную популяцию могут включаться и особи других возрастов, но близкие по своему биологическому состоянию к "ядру", составляющему элементарную популяцию. Такие элементарные популяции отмечены у хамсы, кильки, воблы, трески и некоторых других рыб.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'