Предельный и средний возраст разных видов и популяций рыб
Продолжительность жизни рыб весьма различна: от нескольких месяцев у некоторых бычков до почти ста лет у некоторых осетровых*. Продолжительность жизни и максимальные размеры, достигаемые нашими промысловыми рыбами, колеблются от 10 см и возраста 2 года до размеров более 3 м и возраста нескольких десятков лет (табл. 51).
* (Приводимые иногда в литературе случаи поимки щук с метками, проживших якобы несколько сот лет (щука Фридриха Барбароссы, щука царя Алексея Михайловича), основаны на недоразумении.)
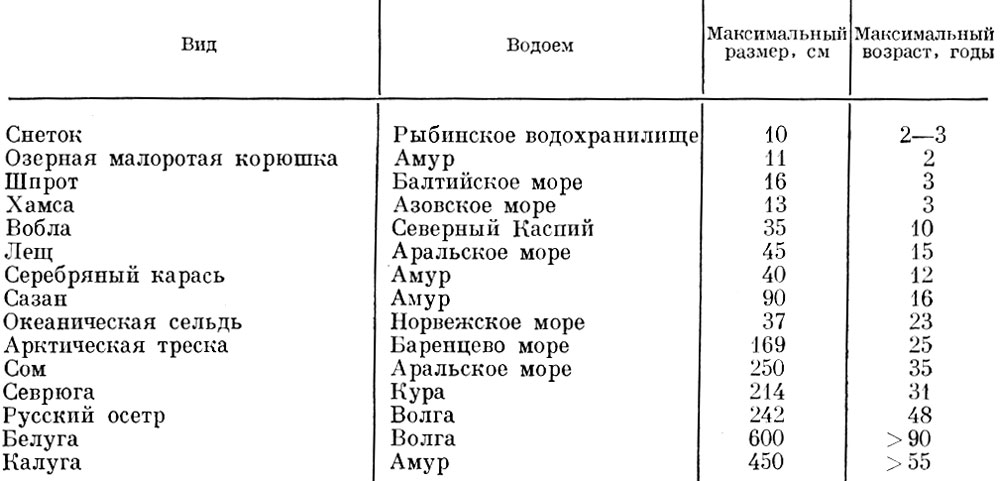
Таблица 51. Обычные предельные размеры и возраст некоторых основных промысловых рыб
Если мы посмотрим, как распределяются виды рыб современной ихтиофауны по предельным размерам и возрасту, то окажется, что наибольшее число видов рыб имеет предельный возраст 5-15 лет и длину 30-50 см (табл. 52). Всего использованы сведения о 177 видах рыб по максимальной длине и о 104 видах рыб по максимальному возрасту.

Таблица 52. Распределение видов рыб по предельному возрасту и предельным размерам (по Beverton a. Holt, 1959; Дрягину, 1934, и нашим данным)
В эту таблицу нами не включены данные о размерах рыб из уловов ископаемого человека. Для некоторых видов они показывают несколько большие величины, чем наблюдаются в ныне живущих популяциях этих рыб. Так, в уловах ископаемого человека обнаружены остатки севрюги, достигавшей длины 270 см и возраста 41 год. Русский осетр соответственно имел 300 см и возраст более 50 лет (Цепкий и Соколов, 1972).
Конечно, это данные ориентировочны, так как охватывают менее 5% современной фауны. Они основаны главным образом на видах рыб умеренных широт северного полушария. В тропических и экваториальных водоемах средняя продолжительность жизни рыб, видимо, несколько меньше. Однако и приведенные данные позволяют сделать некоторые выводы. Около 75% видов рыб имеют продолжительность жизни от 2 до 20 лет, около 60% - от 5 до 20 лет, менее 10% видов живут более 30 лет и около 5% живут менее двух лет. Правда, последняя цифра, вероятно, несколько занижена, так как литературных данных о возрасте рыб с коротким жизненным циклом несколько меньше, чем о рыбах, достигающих среднего свойственного рыбам возраста. Самые крупные рыбы с наибольшей продолжительностью жизни - это, как правило, крупные хищники, обычно с коротким периодом интенсивного нагула. Основное питание обычно осуществляется во время подхода жертв - проходных или полупроходных рыб или концентрации на местах нереста, или перед нерестом. Так питаются белуга, калуга, сом. О крупных морских хищниках мне надежных данных найти не удалось. Исключение в этой группе представляют крупнейшие планктоноядные акулы Cetorhinus maximus Gunner, Rhincodon typus Smith, но и они, видимо, частично питаются и рыбной пищей.
К группе рыб, достигающих средних размеров (до 1 м и немного более) и возраста до 30 лет, относятся главным образом бентофаги, частично растительноядные рыбы и хищники-рыбоеды. Планктоноядных рыб среди этой размерно-возрастной группы сравнительно мало. Среди мелких рыб с коротким жизненным циклом преобладают планктонофаги и питающиеся мелким бентосом рыбы. Хищников среди мелких рыб с коротким жизненным циклом почти нет*.
* (Мы говорим во всех случаях о питании взрослых рыб, а не молоди.)
И у разных популяций одного и того же вида предельный возраст и максимальные размеры могут быть весьма различными, отражая приспособленность популяции к тем условиям, в которых она существует (табл. 53-58). Почти такое же разнообразие наблюдается у основного хищника, питающегося сельдью, - у трески (табл. 54).
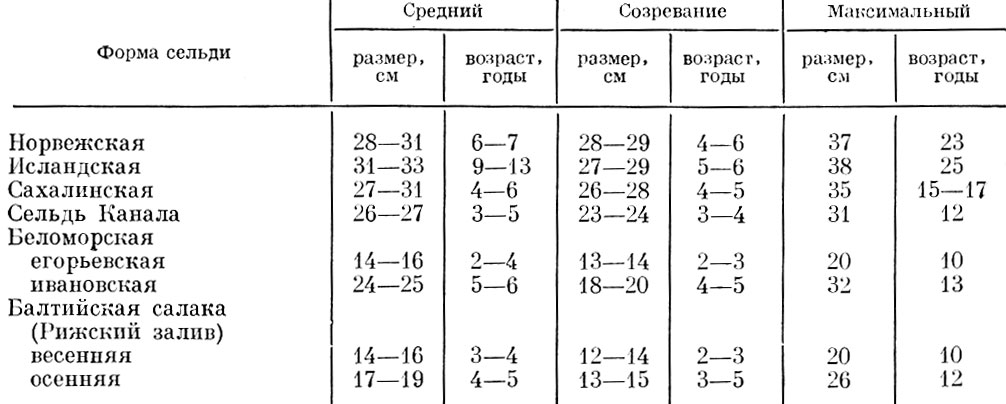
Таблица 53. Средний и максимальный размеры и возраст различных форм океанической сельди Clupea Harpngus L
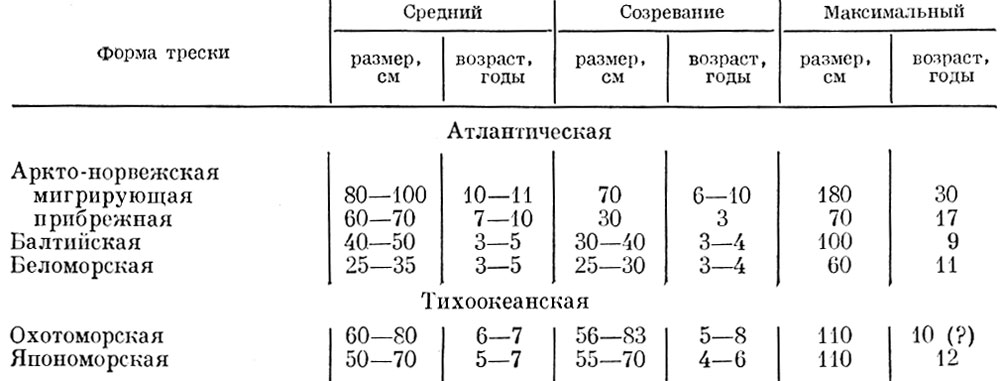
Таблица 54. Средний и максимальный размеры и возраст различных форм трески (по Rollefsen, 1954; Ruud, 1939; Hylen, 1964; Маслову, 1960; Моисееву, 1953 и др.)
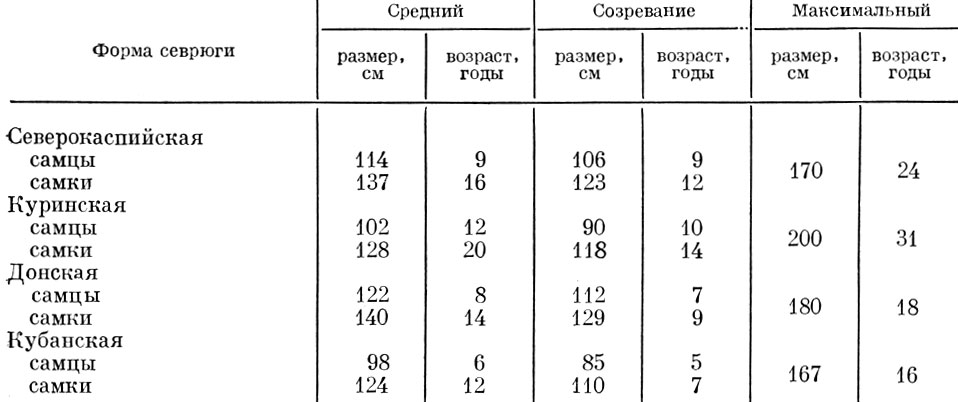
Таблица 55. Средний и максимальный размеры (до конца средних лучей хвостового плавника) и возраст различных стад севрюги (по материалам Н. И. Чугуновой)
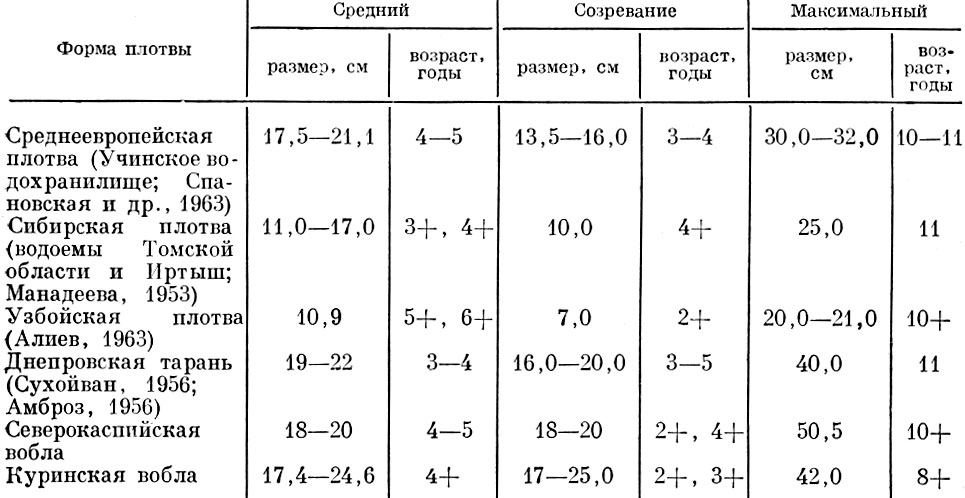
Таблица 56. Средний и максимальный размеры и возраст различных форм плотвы
Как видно из приведенных данных, у разных форм трески, как и у сельди, имеются весьма существенные различия в структуре стада. Существенные различия в структуре стада наблюдаются также у проходных и пресноводных рыб (табл. 55).
Так, у севрюги даже в пределах одного бассейна наблюдаются существенные различия в структуре стад. Столь же существенны различия в структуре стада и у различных форм обыкновенной плотвы Rutilus rutilus L. (табл. 56).
Существенные различия наблюдаются не только у рыб с большой и средней продолжительностью жизни, но и у рыб с коротким жизненным циклом. Правда, у этих рыб различия оказываются несколько меньше (табл. 57 и 58).

Таблица 57. Средний и максимальный размеры и возраст различных форм шпрота Sprattus Sprattus
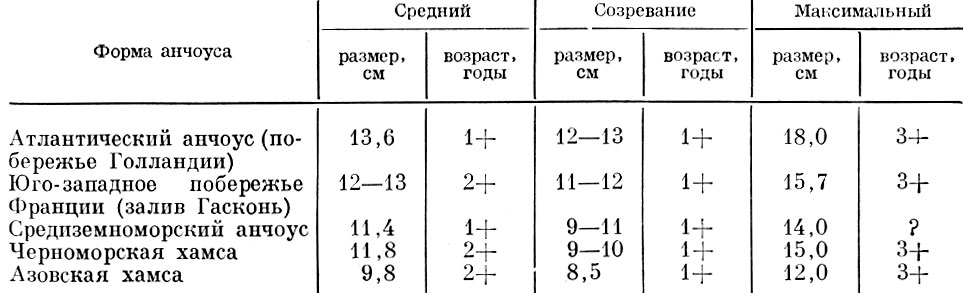
Таблица 58. Средний и максимальный размеры и возраст различных форм обыкновенного анчоуса Engraulis Encrassicholus
Конечно, приведенные данные в ряде случаев лишь ориентировочны, но все же они позволяют отметить некоторые закономерности. Среди всех морских рыб более южные (в северном полушарии) популяции оказываются обычно более мелкими, с меньшим предельным и средним возрастом. Все рассмотренные южные популяции рыб оказываются и раньше созревающими.
Подобные различия в структуре более южных (более раннее созревание, меньшие размеры, меньший средний и предельный возраст) и более северных популяций, несомненно, связаны в первую очередь с разной интенсивностью воздействия хищников и являются приспособлением, обеспечивающим более интенсивное воспроизводство.
Прибрежные жилые популяции даже в одном и том же водоеме обычно характеризуются более коротким жизненным циклом и меньшими размерами по сравнению с мигрирующими.
В данном случае разница в структуре жилых и мигрирующих популяций связана в первую очередь с разницей в обеспеченности пищей. О том, что во многих случаях это не связано со степенью воздействия хищников, свидетельствует хотя бы сравнение беломорской и аркто-норвежской трески. Несомненно, что степень воздействия хищников на беломорскую треску много меньше, чем аркто-норвежскую. Разница же в обеспеченности пищей у этих популяций выражена очень резко.
Значительно более сложная картина наблюдается у пресноводных и проходных рыб.
У разных популяций севрюги различия в структуре популяции так же, как, видимо, и у большинства других осетровых, связаны в первую очередь с разницей в обеспеченности пищей. Значительно более сложная картина наблюдается у кумжи Salmo trutta L. Речные популяции этой рыбы, отличающиеся коротким жизненным циклом и ранним половым созреванием, а также речные части популяций проходных форм (самцы) характеризуются коротким жизненным циклом и часто (но не всегда) более ранней зрелостью. Интересно, что эти популяции по характеру питания отличаются одна от другой так же, как разные виды рыб с коротким и длинным жизненным циклом. Проходные крупные рыбы, как правило, хищники, а жилые мелкие питаются беспозвоночными.
У плотвы структура популяции у разных форм очень разнообразна. У пресноводных форм, питающихся растительностью и мелкими беспозвоночными, наблюдается то же явление, что и у морских рыб: более южные популяции созревают в более раннем возрасте, иногда более мелких размеров, чем северные, и продолжительность жизни у них меньше. Это различие так же, как и у морских форм, связано с приспособленностью южных форм к более интенсивному воздействию хищников. Полупроходные формы, питающиеся моллюсками, достигают половой зрелости раньше, чем пресноводные формы, но при тех же размерах, что и пресноводные формы тех же широт, как это наблюдается у воблы Штеттинской бухты и у распространенных в этих же широтах пресноводных форм плотвы (Желтенкова, 1949).
У южных полупроходных форм плотвы (каспийская и аральская вобла) половое созревание обычно наступает в том же возрасте, что и у пресноводных форм и у прибрежных жилых, так называемых камышовых форм (Гладков, 1933), однако размеры, при которых полупроходные популяции достигают половой зрелости, оказываются значительно крупнее, и обычно возрастная структура стада у полупроходных сложнее, т. е. включает больше возрастных групп.
Такие различия в структуре популяций у северных полупроходных и жилых форм плотвы, несомненно, связаны с приспособленностью к разной обеспеченности пищей. Жилые северные формы живут при более низкой обеспеченности пищей, чем полупроходные. Интенсивность воздействия хищников и на жилые, и на полупроходные северные популяции меньше, чем на южные. Южные жилые популяции достигают половой зрелости при меньших размерах, что связано с приспособлением к высокому прессу хищников при низкой кормовой базе. У южных полупроходных форм размеры, при которых наступает половое созревание, сохраняются близкими к тому, что наблюдается у северных форм. У полупроходных форм, живущих при высокой обеспеченности пищей, снижение интенсивности воздействия хищников частично достигается посредством более быстрого роста.
Близкая картина изменчивости структуры популяций к описанной у разных форм плотвы наблюдается и у некоторых других пресноводных рыб, например у леща. У южных стад прибрежных жилых и некоторых пресноводных форм этого вида половое созревание также происходит при меньших размерах, а полупроходные стада созревают при тех же размерах, что северные формы, но в более раннем возрасте. Приспособительное значение этого явления у леща, несомненно, то же, что и у плотвы.
Впервые в отечественной литературе механизм образования карликовых форм рассмотрен В. В. Васнецовым (1947) на примере тугорослых популяций обыкновенного карася Carassius carassius (L.). Альм (Aim, 1959) связывает образование карликовых форм с изменением качественного состава пищи. К этой же группе явлений относится образование карликовых озерных и речных форм у многих лососевых родов, как Salmo, так и Oncorhynchus (Koshin a. Protassov, 1959; Моисеев, 1957; Смирнов, 1959 и др.) и образование жилых форм корюшек как рода Osmerus, так и рода Hypomesus.
Среди карповых, кроме упомянутого выше образования тугорослых карликовых форм у карася, плотвы и леща, образуются раносозревающие тугорослые формы у сазана и других видов.
Карликовые формы могут периодически возникать от крупных проходных форм при определенных условиях и в свою очередь давать начало крупным проходным особям. Это хорошо прослежено Е. М. Крохиным (1967) для красной оз. Дальнего на Камчатке. Чем выше биомасса кормов в озере, где нагуливается молодь красной, или чем меньше численность поколения, тем больше молоди остается в озере и не уходит в море. То есть чем выше обеспеченность пищей в нерестовом водоеме, тем больший процент молоди остается в нем и превращается в карликовую форму. Интересно, что при этом происходят и некоторые изменения в кариотипе (Черненко, 1969).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'