Закономерности роста рыб
Рост - это количественная сторона развития. Рост рыбы, как и любого другого организма - это увеличение его размеров и массы тела. Рост рыбы - это один из важнейших механизмов, при помощи которого отдельная особь и популяция в целом автоматически реагируют на изменение обеспеченности пищей перестройкой темпа своего размножения и интенсивности и характера потребления кормов, а также и процесса старения (Васнецов, 1947, 1953а).
Рост любого организма, в том числе и рыбы, теснейшим образом связан с обеспеченностью пищей и может изменяться в результате изменения кормовой базы популяции, т.е. увеличения или уменьшения численности биомассы или доступности кормовых организмов вследствие удлинения или сокращения сезона нагула; это, например, бывает в годы потепления у относительно тепловодных видов, когда их обеспеченность пищей повышается, и в годы похолодания у относительно холодноводных. Обеспеченность пищей может увеличиваться и рост рыб ускоряться в результате сокращения численности популяций других организмов, включая и другие виды рыб-потребителей тех же кормов. Наконец, изменение обеспеченности пищей теснейшим образом связано с изменением численности особей данного вида, что приводит к изменению количества корма, приходящегося на одну особь. Изменение численности особей данного вида отражается на обеспеченности пищей особей тех размеров и возраста, которые питаются oсходной пищей. Во всех этих случаях изменение темпа роста приводит к изменению сроков полового созревания (а также количества и качества половых продуктов), вследствие чего меняются структура половозрелой части популяции и интенсивность ее воспроизводства.
Процесс роста в известных пределах специфичен для разных видов рыб. Он является, как и другие видовые признаки, приспособлением, обеспечивающим единство вида и среды.
Как хорошо известно (Васнецов, 1947, 1953а), быстрый рост и крупные размеры позволяют рыбе меньше подвергаться воздействию хищников, лучше защищают ее от врагов. С другой стороны, крупные размеры позволяют рыбе питаться крупной пищей, т. е. расширяют ее кормовую базу. Но в то же время быстрый рост и достижение крупных размеров связаны с потребностью в значительном количестве кормов, а это в свою очередь ограничивает численность популяции крупных рыб.
Для роста и существования мелких рыб требуется сравнительно небольшое количество корма, их популяция может достигать сравнительно высокой численности при ограниченной кормовой базе. Однако малые размеры делают мелких, медленно растущих рыб более доступными для хищников. Мелких рыб хищники выедают значительно интенсивнее, чем крупных, и это связано с выработкой у них более интенсивного темпа воспроизводства.
Одной из основных характерных черт процесса развития и роста как количественной стороны развития всего живого является то, что индивидуальное, а также историческое развитие организмов - это прерывистая непрерывность. У всех видов живых организмов, как мы видели, процесс развития от зачатия до смерти распадается на отдельные этапы, характеризующиеся своими внутренними противоречиями, своими специфическими связями со средой, своими ведущими отношениями (Васнецов, 1948, 1953; Крыжановский, 1949, 1950). Характер роста специфичен для каждого этапа развития. Переход с этапа на этап происходит у каждого вида рыбы за сравнительно немногими исключениями примерно при одном и том же размере (Васнецов, 19536). Характер роста определяется особенностью тех условий, приспособлением к которым рост является, и специфичностью тех физиологических механизмов, через которые он реализуется.
Представление о росте как только о постепенном процессе и связанные с таким представлением попытки построения единого для всего онтогенеза уравнения роста являются, как мне кажется, бесплодными. Я не могу согласиться с Бевертоном и Холтом (Beverton a. Holt, 1957), что возможна единая "работающая" формула роста, которая удовлетворительно характеризовала бы рост большинства видов рыб на всех этапах развития. Ни в коей мере не отвечает этим целям и формула Берталанффи (Bertalanffy, 1934, 1938 и др.), которую пытаются использовать Бевертон и Холт (1957). Попытки вывести единую формулу роста, характеризующую рост всех рыб, а может быть и вообще всех организмов, в основе которой лежит уравнение, отражающее ход определенных химических реакций, мне представляются и методологически неправильными. Это приводит к забвению специфики живого, к забвению того, что рост - это приспособительный процесс. В конечном итоге это порождает попытки, вроде попытки Рудольфа Карнапа (по М. Корнфорту, 1957), найти единую формулу, характеризующую все физические и биологические процессы. Взгляды Карнапа Берталанффи в значительной мере и развивает. Как отечественные (Шмальгаузен, 1928, 1935; Васнецов, 1934, 1953а), так и зарубежные исследователи (Le Gren, 1958; Ricker, 1958; Parker a. Larkin, 1959 и др.) показали, что кривая роста распадается на ряд отрезков, подчиняющихся своим специфическим закономерностям.
И. И. Шмальгаузен (1935), вычисляя константу роста, установил, что она специфична для отдельных отрезков индивидуального развития. То же установлено и В. В. Васнецовым (1934 и др.) путем анализа изменений характеристики роста*. Паркер и Ларкин (1959) пришли к выводу, что процесс роста может быть выражен в общем виде уравнением (в данном случае для роста массы)
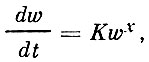 (5)
(5)где:
ω - масса;
t - время.
* (Характеристика роста, по В. В. Васнецову (1934), вычисляется по формуле)
| Х = | lgl2 - lgl1 | l1 |
| 0,4343 (T1 - T1) |
где:
l1 и l2- длина;
Т1 - Т2- промежуток времени.
Коэффициенты К и х специфичны для каждого естественного отрезка онтогенеза, причем К, по мнению этих авторов, отражает специфику экологических отношений, определяющих рост, а х: - соответственно специфику физиологических процессов.
Конечно, такое разделение на экологические и физиологические процессы условно; изменения этих коэффициентов происходят за короткий промежуток времени, когда особь достигает определенного размера, специфичного для данного вида и данного этапа. Эти авторы отмечают, что у отдельных видов в некоторых случаях, когда популяция "меняет свою экологическую нишу", установившийся пороговый размер может меняться.
Уравнение Паркера и Ларкина, как это указывают Полохеймо и Дикки (Poloheimo a. Dickie, 1965), отражает процесс роста при постоянной эффективности использования кормов. В случае же непостоянства "оплаты корма", что, как известно, свойственно всем рыбам, формула требует известной корректировки.
В. Л. Брюзгин (I960,1968,1969) правильно отмечает, что законы роста рыб в длину отличаются от математического закона органического роста, подчиняющегося правилу сложных процентов. Ряд его критических замечаний в отношении предложенных И. И. Шмальгаузеном (1935) формулы "удельной скорости роста" и В. В. Васнецовым (1934) характеристики роста, несомненно, правильны, однако, как мне представляется, характеристика роста вполне применима для выяснения периодики и ряда других вопросов специфики роста (см., например, Сиверцов, 1963).
Естественно, что поскольку рост рыбы - это ее видовое приспособление, то на изменения обеспеченности пищей рыба приспособительно реагирует изменением темпа роста.
Как показано многими исследованиями (Васнецов, 1934; Монастырский, 1940; Aim, 1959 и др.), время наступления половой зрелости у большинства видов рыб обычно связано с достижением определенных размеров, а не возраста, т. е. физиологический возраст далеко не всегда отражает хронологический. У леща, например, обычный размер достижения половой зрелости 27-28 см (Васнецов, 1934,), у жилых форм плотвы - около 12 см.
Однако из этого общего правила имеются многочисленные исключения. Это в первую очередь карликовые формы многих видов рыб (Кошелев, 1971). Это же наблюдается у разных по характеру питания и темпу роста форм гольцов Salvelinus alpinus (L.) и сигов Coregonus lavaretus (L.) (Савваитова, 1966; Решетников, 1966), а также у проходных и жилых форм лососей и кумжи бассейнов Атлантического и Тихого океанов.
Но и в пределах карликовых, медленно растущих и рано созревающих популяций, при меньших размерах достижения половой зрелости снова устанавливается связь ее с размерами, а не с возрастом (Лапин и Юровицкий, 1959). То, что время наступления половой зрелости, а тем самым и темп воспроизводства популяции в значительной степени зависят от скорости роста рыб, делает понятным, что у рыб, не достигших половой зрелости, значительная часть поступающих в организм пищевых ресурсов идет на увеличение их размеров. Рост рыб в длину до достижения ими половой зрелости в значительно большей степени зависит непосредственно от обеспеченности пищей в данное время, чем после достижения половой зрелости. Быстрый линейный рост до достижения половой зрелости обеспечивает рыбе, с одной стороны, выход из-под воздействия хищников в более молодом возрасте, т. е. снижает интенсивность смертности, и, с другой стороны, достижение половой зрелости в более молодом возрасте ускоряет темп воспроизводства популяции. К сожалению, специфика роста на отдельных этапах развития до достижения половой зрелости почти еще не изучена. Можно только отметить, что по мере перехода с этапа на этап у рыб и до достижения половой зрелости снижается интенсивность потребления корма. Ход этого снижения, естественно, различен у разных видов рыб. Так, и у плотвы, и у леща, и у окуня по мере роста интенсивность потребления пищи резко падает (табл. 45).
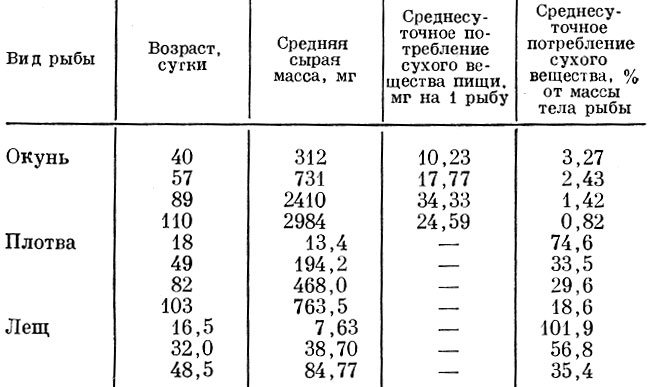
Таблица 45. Величина потребления пищи молодью некоторых пресноводных рыб (по Карзинкину, 1952)
У всех сравниваемых в табл. 45 рыб интенсивность потребления пищи по мере роста быстро падает. Естественно, что при питании разными кормами суточные рационы молоди сильно меняются, как это хорошо показано на большом материале Г. С. Карзинкиным (1952).
По мере роста рыбы кормовой коэффициент у нее становится все выше (Карзинкин, 1952) и очень быстро снижается процент продуцирующего корма, используемого на рост по отношению к поддерживающему корму. Питание одного и того же вида рыбы разными кормами (например, молоди осетра циклопами и малощетинковыми червями) характеризуется разными показателями, но в обоих случаях по мере роста рыбы наблюдается снижение процента азота пищи, используемого на рост (Карзинкин, 1953). Так, у ушастого окуня Lepomis megalotis массой 10 г на рост используется 33% протеина пищи, а у рыб массой 105 г - только 5% потребленного протеина (Gerking, 1952). То же самое на ряде видов рыб показано Полохеймо и Дикки (Poloheimo a. Dickie, 1966a).
Меняются кормовой коэффициент и соотношение поддерживающего" и продуцирующего кормов и у рыбы, достигшей половой зрелости. У старых рыб относительная величина продуцирующего корма по отношению к поддерживающему становится ничтожной. То, что у рыб, не достигших половой зрелости, основная часть поступающих в организм кормов тратится на линейный рост, связано с большей зависимостью роста на первых годах жизни от обеспеченности пищей и в первую очередь от кормовой базы водоемов. Как показал В. В. Васнецов (1934), линейная характеристика роста оказывается наиболее изменчивой до достижения рыбами половой зрелости. Из приведенных данных о леще хорошо видно, что самые сильные различия в характеристике роста наблюдаются у неполовозрелых рыб. После достижения рыбами половой зрелости величина линейной характеристики, их роста как в малокормных, так и в высококормных водоемах становится более или менее близкой (рис. 16).
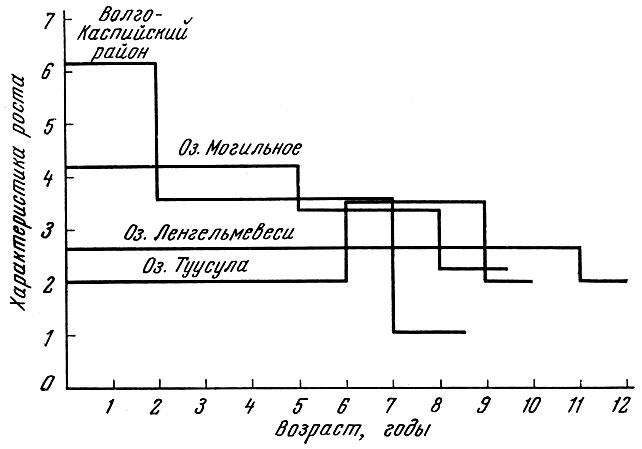
Рис. 16. Характеристика линейного роста леща в различных водоемах (по В. В. Васнецову)
В водоемах, где популяция рыб находится в условиях низкой обеспеченности пищей, например популяция леща финских озер, линейная характеристика роста до достижения рыбами половой зрелости оказывается ниже, чем после достижения половой зрелости. У рыб, не достигших половой зрелости, период линейного роста в течение года продолжительнее, чем у половозрелых. Рост неполовозрелых рыб замедляется лишь в особо неблагоприятные периоды времени. Так, например, годовики форели быстрее всего растут весной и осенью, но рост продолжается в течение всего года (Swift, 1961). У половозрелых рыб - трехгодовиков - сохраняется такая же ритмика, но в отличие от молоди скорость роста резко снижается в июле и августе (Swift, 1955). Неполовозрелый аральский лещ интенсивно растет в длину весь период года с температурами воды выше 14,5° С, т. е. с апреля по октябрь (Константинова, 1958). Так как резервных веществ в организме неполовозрелой рыбы не накапливается, то неполовозрелый лещ питается круглый год, однако зимой он растет очень медленно вследствие того, что резервных веществ в теле неполовозрелых рыб содержится очень мало и малейшее изменение в обеспеченности пищей у неполовозрелых рыб тотчас сказывается на скорости их роста.
Естественно, что рост рыбы определяется не только теми условиями, в которых живет данная рыба, но и теми условиями, в каких жили ее родители, т. е. ее наследственными свойствами. Как было показано выше, от условий нагула родительского стада во многом зависят запас желтка в икринке, ее жирность и разноразмерность выходящих из оболочки свободных эмбрионов. Потомство от особей разного возраста, полученное в одной и той же обстановке, растет по-разному. Это хорошо видно на примере молоди карпа, полученной от производителей разного возраста (Сиверцов, 1963, 1963а). Разная скорость роста сохраняется и у рыб на втором году жизни. В потомстве одной и той же особи, выращиваемом в сходных условиях, как это показано А. В. Морозовым (1951) на примере потомства, полученного от одной самки осетра, одни мальки росли лучше, другие хуже. Сходные данные получены также Б. С.Матвеевым (1951) и С. В. Емельяновым (1961), а для окуня - Л. К. Ильиной (1970).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'