4. Почти совершенное сообщество
Из всех новых форм животных, открытых в гидротермальных сообществах, большой трубчатый червь, названный в 1981 году Riftia pachyptila, является сегодня наиболее известным. Его длина может превышать 1,5 м, а ярко-красный, киноварный цвет его султана, контрастирующего с перламутрово-белой окраской трубки, придают подводным фотографиям с его изображением неоспоримую эстетичность. Внутренее его строение также удивительно: этот червь не имеет ни рта, ни пищеварительной системы, ни анального отверстия!* Наконец, по биомассе этот червь во много раз превосходит все другие виды гидротермальных сообществ.
* (В 1988 году установлено, что личинки и очень ранняя молодь вестиментифер имеют рот, кишку и анальное отверстие, которые исчезают вскоре после установления симбиоза с бактериями. - Прим. ред.)
Для специалистов, однако, рифтия не является совершенной незнакомкой. Она входит в тип беспозвоночных, открытый в 1914 году во время экспедиции на голландском океанографическом судне "Сибога" в воды Индонезийского архипелага. Описавший червя французский зоолог Коллери, пораженный характерным видом его трубок, окрестил червя Siboglinum (от Siboga - название судна и linum - нить по-латыни) и не колеблясь отнес его к новому типу беспозвоночных. Коллери отметил полное отсутствие у червя пищеварительной системы и, пытаясь объяснить это, предположил, что Siboglinum - колониальное животное, у которого функции питания и размножения могут выполняться разными индивидуумами, как это часто имеет место у гидрозоев: отсутствие в собранных материалах других индивидуумов, ответственных за питание колонии, было, однако, трудно понять.
Эти крохотные червеобразные животные живут в хитиновых трубках правильной цилиндрической формы диаметром примерно 1 мм.
Лет через тридцать после их открытия швед Йохансон объединяет Siboglinum с другой, весьма отличной формой из Охотского моря, Lamellisabella в новый класс морских беспозвоночных - погонофоры (от греческого "кто носит бороду", другими словами, бородатые...), быстро возведенный зоологами в ранг подтипа. Открытия множатся, замечено, что представители "бородатых", считавшиеся обитателями исключительно больших глубин, могут жить и на глубине нескольких десятков метров и встречаются гораздо чаще, чем предполагалось ранее. Но все эти погонофоры невелики по размерам и все лишены пищеварительной системы. Первую гипотезу, разрешающую эту загадку, выдвинул советский ученый А. В. Иванов. Согласно его гипотезе эпидермальные клетки передних щупалец погонофор способны вылавливать мелкие органические частицы путем фагоцитоза. Но поскольку наблюдать фагоцитоз не удалось, Иванов позднее выдвинул другую гипотезу: собранные султаном щупалец частицы перевариваются вне тела животного ферментами, выделяемыми в наружную среду, а мелкие молекулы, образующиеся в ходе ферментативного разложения частиц, затем абсорбируются проницаемой клеточной мембраной. Второй гипотезе повезло не больше, чем первой, и она так и не была доказана.
В 1966 году на глубине 1125 м у побережья Калифорнии открыты погонофоры, которые зоолог Вебб назвал Lamellibrachia barhami. Они отличаются от всех других наличием особого вестиментального участка тела, на котором на спинной стороне открываются парные половые отверстия. Этот участок расположен между щупальцевой короной и туловищем и снабжен двумя складками, или крыльями, благодаря которым между вестиментумом и стенкой трубки червя образуется дорсальная камера. Чтобы отметить эту особенность, а также другие, не менее важные черты строения, в типе погонофор создаются новый отряд - вестиментиферы и новый класс - афренуляты. Характерная для типа "бородатых" сегментированная и снабженная хитиновыми щетинками задняя часть тела позволяет сблизить их с кольчатыми червями. Но по-прежнему неизвестно, как и чем питаются эти странные животные.
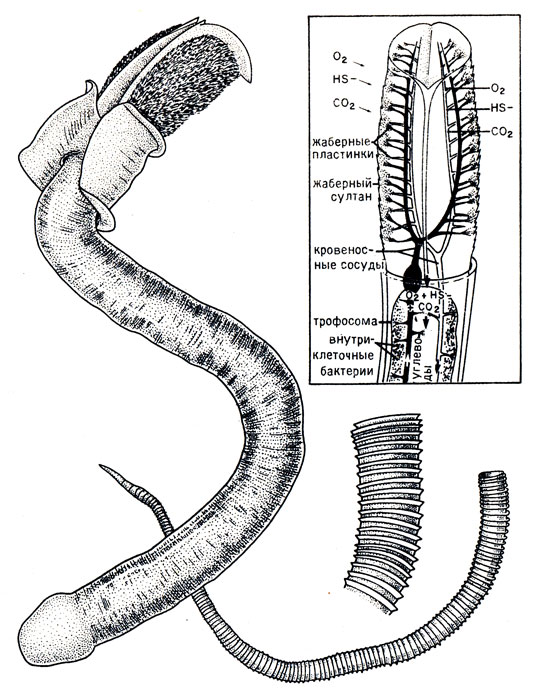
Рис. 17. Гигантская погонофора-вестиментифера Rittia pachyptila, извлеченная из трубки, и фрагменты трубки рифтии и кольчатой трубки вестиментиферы Tevnia jerichonana. На врезке - циркуляция крови в султане щупалец и газообмен между морской средой и эндосимбиотическими бактериями в трофосоме
Вот так в общих чертах обстояло дело, когда в 1977 году на излиянии на Галапагосском рифте был открыт новый вид Riftia pachyptila. Изучением первых собранных образцов занимается американский зоолог Джоунс; менее чем через четыре года после первых находок опубликовано описание вида. Это действительно вестиментифера, но столь сильно отличающаяся от Lamellibrachia, что для нее приходится создать особое семейсто - Riftiidae. Одновременно Джоунс устанавливает, что туловище, занимающее около 3/4 длины тела, заполнено тканью, образованной дольками, обильно снабженными кровеносными сосудами, - трофосомой; эта ткань буквально насыщена округлыми бактериями диаметром около 3-5 мкм. Трофосома различима даже у самых молодых изученных особей (длиной 1,44 мм) и содержит во множестве кристаллы серы.
Спустя несколько месяцев, в июне 1981 года, в одном из номеров высокоавторитетного журнала "Сайнс" ("Наука") - органа Американской ассоциации развития науки - появляется целая серия статей, посвященных рифтиям. В них сообщается множество результатов, подтверждающих, что рифтия получает питание от бактерий, содержащихся в трофосоме, или потребляет их метаболиты. Хотя доказательства еще далеко не исчерпывающие, приведенные данные в целом весьма определенно ориентированы в эту сторону.
Первый комплекс данных о содержащихся в трофосоме бактериях получен с помощью электронного микроскопа и свидетельствует о том, что это сферические грамотрицательные бактерии, мембрана которых содержит липополисахариды. Полученные изображения весьма сходны с ранее опубликованными фотографиями бактерий из трофосомы Lamellibrachia, первой известной вестиментиферы (тогда Вебб чисто гипотетически отвел трофосоме роль органа-накопителя продуктов, необходимых для сперматогенеза, или органа детоксикации, не имея каких бы то ни было доказательств).
Интереснее и существеннее результаты, полученные биохимиком Фельбеком. Прежде всего, ткани трофосомы имеют повышенную активность многих ферментов, входящих в цикл окисления - восстановления восстановленных соединений серы (например, тиосульфат-редуктазы, или роданезы, и аденозинтрифосфатсульфурилазы). Это доказывает, что сферические бактерии из трофосомы рифтии участвуют в цикле серы (в частности, окислении сероводорода). Другие опыты показали, что бактерии трофосомы содержат также характерные ферменты фиксации СО2 в цикле, называемом С3-циклом или циклом Кальвина-Бенсона, широко распространенном у высших растений. Первое органическое соединение, образованное из атмосферного СО2, содержит три атома углерода и его синтез идет с участием фермента рибулозо-1,5-бифосфаткарбоксилазы (более известного биохимикам под названием RuBP-аза). Эта RuBp-аза содержится в трофосоме рифтии, но полностью отсутствует в мышцах червя. То же относится и к другому ферменту, рибулозо-5-фосфаткиназе (или Ru5P-киназe). Таким образом, можно предположить, что рифтия, благодаря заключенным в ее трофосоме бактериям, способна кормиться либо полностью как автотрофный организм, либо частично, за счет энергии окисления сероводорода, способствующего восстановлению СО2 в органические соединения.
Еще один, последний результат касается измерения отношения двух стабильных изотопов углерода 13С и 12С в трофосоме и в остальном теле рифтии. Известно, что отношение двух природных изотопов углерода практически не меняется при переходе с одного уровня пищевой пирамиды на другой, так как оно зависит от величины этого отношения в источнике минерального углерода и изначально определяется способом фиксации СО2. Это свойство позволяет использовать изменение отношения двух природных изотопов элемента, обычно присутствующего в цикле органического вещества, в качестве метки для пищевой цепи.
Измерения соотношения изотопов углерода (13С / 12С), проведенные Pау в трофосоме и в мышцах рифтии, дали одинаковые значения - примерно - 11% (эта величина выражает отклонение между соотношением изотопов в исследуемой пробе и в стандартном образце, например в воздухе, вычисленное по формуле: отношение в исследуемой пробе, поделенное на отношение в стандартном образце, минус 1, выраженное в промилле). Тот факт, что и в трофосоме и в мышцах было получено одно и то же значение, указывает, что эти две ткани непосредственно связаны трофически, иными словами, что бактерии или их метаболиты служат пищей для погонофор. Абсолютное значение отношения интерпретировать труднее. В самом деле, у морских животных, которые питаются органическими частицами, произведенными одноклеточными водорослями фитопланктона, значения отношения колеблются от -15 до -18%. По сравнению с этими величинами, относительно высокие значения, полученные для тканей рифтии, могут быть объяснены тремя причинами: либо тем, что их пищу составляют остатки растений, функционирующих по циклу С4, что представляется маловероятным; либо тем, что бактериальный хемосинтез идет в условиях недостатка СО2, вследствие чего ферменты цикла Кальвина оказываются менее селективными по отношению к двум изотопам углерода; либо тем, что для переноса СО2 бактерии, подобно некоторым зеленым растениям, используют четырехуглеродный сахар малат.
Фельбек продемонстрировал правильность последнего объяснения: он вносил радиоактивно меченую СО2 в рифтий, через несколько часов после сбора помещенных в охлажденный до 8 °С аквариум под давлением 110 атмосфер, и после нескольких часов инкубации обнаруживал в крови рифтий меченые молекулы малата и сукцината. Таким образом, СО2 поступает до трофосомы, где высвобождается путем декарбоксилирования.
Наконец, изучение крови рифтий показывает, что своим цветом она обязана повышенному содержанию гемоглобина (в крови рифтий нет оформленных элементов, как у большинства беспозвоночных) и что она обладает повышенной способностью переносить кислород (до 11 мл кислорода на 100 мл крови). Подобное свойство может объясняться большой потребностью в кислороде для бактериального хемотрофного метаболизма, которая добавляется к потребности в кислороде для дыхания.
Эти результаты еще не позволяют утверждать, что рифтия питается за счет хемосинтезирующих бактерий, живущих в ее трофосоме: надо еще объяснить, каким образом сероводород, извлекаемый султаном щупалец из морской воды, доносится до бактерий, которые его окисляют, откуда берется азот, необходимый для построения белков, и как осуществляется перенос бактериального органического вещества к тканям погонофор.
Тем временем открытие Rittia pachyptila стимулировало исследования классических погонофор. Шаг за шагом было экспериментально показано, что поглощения растворенных в морской воде органических веществ, таких, как аминокислоты, при их естественных концентрациях недостаточно для удовлетворения метаболических нужд даже такого маленького червя, как Siboglinum fjordicum; затем в 1981 году - что и эти организмы, как и большие вестиментиферы, содержат хемосинтезирующие внутриклеточные бактерии (были исследованы три вида Siboglinum и один вид Oligobrachia). Отношение изотопов 13С / 12С очень низкое (-45%, что позволяет предполагать использование метана. Наиболее важная сторона этого исследования заключается в том, что оно показало: связь между хемосинтезирующими бактериями и погонофорами отнюдь не ограничивается только вестиментиферами, классические мелкие погонофоры также получают по меньшей мере часть пищи от хемосинтезирующих бактерий, содержащихся в их клетках.
Все это время не прекращались исследования рифтий, которая становится настоящим лабораторным животным. Перенос сульфидов кровью в принципе сталкивается с многочисленными трудностями: сульфиды образуют с гемоглобином стабильные соединения, которые не могут связывать кислород; более того, в присутствии этого газа они спонтанно окисляются и не могут быть далее использованы бактериями; наконец, они сильно токсичны и подавляют внутриклеточные ферментативные реакции дыхания. В 1983 году две американские исследовательские группы одновременно выявили в крови рифтий белок с повышенной молекулярной массой, способный соединяться с очень большим количеством сульфидов и таким образом защищать гемоглобин крови и внутриклеточные дыхательные ферменты от вредных воздействий. Благодаря этому белку кровь рифтий способна накапливать большие количества сероводорода на уровне султана щупалец и транспортировать его до трофосомы. Более поздние работы показывают, что этот любопытный белок является не чем иным, как гемоглобином, который может, таким образом, обеспечивать перенос кислорода и сульфидов на двух разных участках: кислорода - на геме, а сульфидов - на глобине. Действительно, количество фиксированных сульфидов, как оказалось, прямо зависит от количества гема, и, кроме того, к тому времени было уже известно, что молекулярная масса этого белка весьма близка к молекулярной массе гемоглобина.
Годом позже публикация Боска и Грассе об особо тщательном исследовании трофосомы рифтий в электронном микроскопе приносит целую серию интереснейших результатов. Прежде всего, сферические бактерии трофосомы локализованы внутри клеток ткани трофосомы, что прежде доказано не было. Во-вторых, эти внутриклеточные бактерии присутствуют во всех клетках трофосомы за исключением тех, которые окаймляют кровеносные сосуды, и самых наружных клеток долек. Внимательный анализ клеток, окаймляющих кровеносные сосуды, показывает, что составные элементы крови, представленные структурированными частицами, приклеиваются к наружной поверхности клеточной мембраны и образуют очень характерную сплошную выстилку. Эти частицы поступают в цитоплазму клеток путем пиноцитоза (пиноцитоз - это формирование вакуолей, которые образуют ножку, затем полностью отделяются от клеточной мембраны, на которой они возникли, и проникают в тело клетки). Полученные снимки показывают, что, вероятно, в клетку поступает белок крови, который доносит сероводород до трофосомы. В более удаленных от сосуда клетках появляются бактерии, которые быстро размножаются. Судя по цитоплазматическим включениям клетки-хозяева имеют пониженную метаболическую активность. Этот процесс продолжается в периферических клетках и ведет к гибели бактерий: измененная бактериальная стенка не препятствует более проникновению воды, диаметр бактерии увеличивается более чем на 20%, затем бактерии подвергаются настоящему внутриклеточному пищеварению - бактериальному лизису. Клетка червя, нагруженная продуктами бактериального распада, завершает процесс дегенерации, начинающийся с момента появления первого бактериального симбионта: смешанные продукты бактериального распада и разрушения клетки, которая их содержит (бактериоцита), поступают в кровь погонофоры, - они-то и являются теми веществами, которые обеспечивают выживание и рост червя.
Чтобы быть вполне доказательным, ультраструктурный анализ трофосомы рифтии должен быть подтвержден физиологическими опытами. В частности, сам по себе он не позволяет оценить соотношение органических метаболитов, выделяемых бактериями в ходе их жизнедеятельности, и продуктов посмертного распада бактерий. Американские исследователи провели в 1985 году специальную экспедицию, посвященную физиологическому изучению гидротермальных организмов. Они располагали системой, позволяющей поместить животных, взятых со дна за несколько часов до эксперимента, в замкнутую камеру под давлением, и снабжать их различными необходимыми минеральными соединениями (СО2, сероводород, кислород), непрерывно измеряя определенные физико-химические параметры крови. Нет сомнения, что такие методы помогут нам глубже понять функционирование ассоциации рифтий и связанных с ними бактерий.
Здесь многое еще неясно. Прежде всего, даже если ассоциация с хемосинтезирующими бактериями явно доказана, то значение трансэпидермального питания органическими молекулами, растворенными в морской воде, остается неизвестным: комбинируется ли оно с первым способом питания или же его следует решительно отвергнуть? Во-вторых, цикл азота, необходимого для синтеза белков, еще мало изучен; установлено лишь наличие в трофосоме фермента, способного извлекать азот из молекул нитрата путем денитрификации, - нитратредуктазы. В-третьих, в гидротермальных флюидах присутствуют другие восстановленные соединения: метан и некоторые азотистые вещества. Используются ли они ассоциацией бактерии-рифтия или эта ассоциация специфически тяготеет к сероводороду, что, возможно, и является результатом адаптации червя к гидротермальной среде?
Последний вопрос заставляет задуматься о связях, объединяющих бактерий и погонофор: настоящий ли это симбиоз, как в случае лишайников, которые являются примером постоянного и обязательного симбиоза гриба и водоросли определенных видов, всегда одних и тех же, не способных развиваться один без другого, или же это ассоциация, которая не предполагает такой жесткой специфической связи между двумя организмами? Сейчас еще слишком рано отвечать на этот вопрос. Бактерии, которые содержатся внутри клеток трофосомы, еще не определены, и самые последние наблюдения, похоже, свидетельствуют, что существует, возможно, не меньше двух категорий клеток-хозяев, что предполагает изменчивость на уровне самих штаммов бактерий.
Наконец, пока еще неизвестен механизм передачи бактерий взрослыми червями своим потомкам. Созревающие яйца извлекали из яичников самки рифтии и изучали под электронным микроскопом - оказалось, что они лишены симбиотических бактерий. Однако у трех очень молодых экземпляров рифтии длиной 2 мм Джоунс установил наличие ресничного отверстия в основании султана щупалец, связанного с тонким ресничным канальцем, который проходит через мозг и достигает полости тела. Эта структура исчезает на ранней стадии. Возможно, она представляет собой путь для проникновения бактерий. Но каким образом молодые черви выбирают в окружающей их среде те бактериальные штаммы, с которыми они затем образуют длительную и обязательную ассоциацию, настоящий симбиоз? Или они способны включать в свои клетки бактерии, принадлежащие к разным штаммам? Сейчас невозможно ответить на эти вопросы. Можно лишь подчеркнуть, что у рифтии столько биологических и физиологических адаптации, что гипотеза настоящего симбиоза с вестиментиферами кажется наиболее вероятной.
Наконец, вестиментиферы представляют особенно показательный пример эволюционной модели, которая известна нам пока лишь частично. В самом деле, вновь открытые гидротермальные излияния, например те, что расположены в зоне субдукции или в местах выхода интерстициальных вод, принесли Джоунсу большое количество новых для науки форм. Судите сами: в 1982 году было известно всего три вида вестиментифер, сгруппированных в два семейства, каждое с одним родом: Lamellibrachiidae с видами Lamellibrachia barhami, первой известной вестиментиферой, и L. luymesi, Riftiidae с видом Riftia pachyptila.
В ноябре 1985 года на международном совещании по гидротермальной фауне, состоявшемся в Океанографическом институте в Париже, Джоунс сообщил, что число известных видов вестиментифер утроилось, что они объединяются в пять семейств, не считая некоторых видов, которые пока еще не определены. Шесть новых видов, чьи названия говорят о зонах, где они были собраны, распределены в четыре новых семейства:
- Ridgeiidae (от английского ridge - хребет), с родом Ridgeia, содержащим два вида, собранных на гидротермальных излияниях хребтов Хуан-де-Фука и Эксплорер: Ridgeia piscesae и R. phaeophiale.
- Escarpiidae, с родом Escarpia (от английского escarp - обрыв, склон, эскарп), включающим два вида, собранных в оазисах у подножия Флоридского эскарпа, Escarpia spicata и Е. laminata.
- Tevniidae, с двумя родами: Tevnia (поскольку название Ventia, производное от английского слова vent - выход, источник, уже использовалось в номенклатуре животных, то, чтобы сохранить упоминание о гидротермальной среде, Джоунс прибег к анаграмме) и Oasisia. Род Tevnia с единственным видом Т. jerichonana был собран "Сианой" в 1982 и 1984 годах на 13° с. ш. на Восточно-Тихоокеанском поднятии. Род Oasisia, также включающий единственный вид О. alvinae, был собран "Алвином" на 21° с. ш. на том же поднятии.
В обрабатываемых коллекциях есть еще второй иерихонский червь, добытый на 18° ю. ш. на Восточно-Тихоокеанском поднятии на расстоянии более 3500 км от местонахождения, где был найден Т. jerichonana, и три вида рода Ridgeia.
Создается впечатление, будто эти крупные животные, еще двадцать лет назад не известные науке, способны эволюционировать с такой скоростью, что образуют четко различающиеся виды, роды и семейства, оставаясь строго приуроченными к определенным физико-химическим условиям среды, которые изменяются в узких пределах и, кроме того, значительно различаются в разных гидротермальных излияниях. Следует ожидать, что у всех этих видов будут открыты физиологические механизмы, схожие с теми, которые были обнаружены у Riftia pachyptila, или даже идентичные им.
Наконец, Джоунс предлагает повысить старый подтип погонофор Obturata (иначе говоря, вестиментифер) до ранга типа и числить их под названием Vestimentifera, принимая во внимание следующие их анатомические отличия от собственно погонофор: различия в числе и размере внутренних полостей тела (целомических полостей), конфигурацию вентральной нервной цепочки и наличие особого тканевого листка, мезентерия, в терминальном районе тела. Итак, отныне погонофоры и вестиментиферы составляют два различных типа морских беспозвоночных.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'