Восприятие рыбами слабых электрических полей
В 1917 г. американские ученые Г. Паркер и В. Гензен изучали чувствительность американского сомика к различным раздражителям. Воздействуя на рыб палочками, изготовленными из стекла, дерева и металла, они обнаружили, что сомики чувствуют приближение металлической палочки на расстоянии нескольких сантиметров, а на стеклянную реагируют только при ее прикосновении. Если поверхность соприкосновения металлической палочки с водой составляла 5-6 см2, рыбы уплывали, а при 0,9-2,8 см2 - они подплывали и "клевали" место контакта металла с водой. На электрически изолированные от воды (покрытые слоем парафина) металлические палочки рыбы реагировали так же, как и на стеклянные,- лишь в момент прикосновения. На основании этих наблюдений было сделано предположение, что реакцию сомиков вызывали микротоки, возникающие в результате контакта металла с водой (гальванический эффект).
Дальнейшие эксперименты подтвердили этот вывод. Создавая в воде с помощью электродов и гальванического элемента электрическое поле постоянного тока, Паркер и Гензен обнаружили у рыб такие же реакции, какие вызывали металлические палочки. Токи менее 0,99 мкА вызывали "клев" электродов, больше 1,47 мкА - отрицательную реакцию.
Высокую чувствительность некоторых сомовых рыб к электрическим полям отмечали японские ученые С. Кокубо и К. Узука. При дальнейших исследованиях было установлено, что эти рыбы обладают специализированными электрическими рецепторами.
Иной механизм восприятия электрических полей у рыб, не имеющих электрорецепторов. Электрический ток - универсальный раздражитель нервных и мышечных клеток. Поэтому рыбы реагируют на них при воздействии непосредственно на нервно-мышечные структуры или рецепторы, предназначенные для восприятия неэлектрических раздражителей.
Чувствительность таких рыб к электрическому току исследовалась при изучении их реакций на сильные электрические поля. Ее порог определялся по значению напряжения, при котором наступала первичная двигательная реакция. Однако выяснилось, что этот метод не дает точных данных. Например, у морской лисицы, обладающей чувствительностью в 0,01 мкВ на 1 см, первичная двигательная реакция не совпадает с порогом восприятия электрического поля и проявляется только при напряжении в десятки - сотни вольт. Таким образом, не исключено, что рыбы, не имеющие электрорецепторов, чувствуют более слабые электрические поля.
Следующий этап изучения электрической чувствительности рыб начался с 1958 г., когда Г. Лиссман обнаружил у слабоэлектрических рыб - гимнарха и гимнотуса - особые высокочувствительные электрорецепторы. Стало очевидным, что некоторые рыбы с их помощью ориентируются в окружающей среде. Рыб, имеющих электрорецепторы, стали исследовать многие ученые.
Датские ученые С. Дийкграф и А. Кальмджин в 19G2 г. установили, что высокой чувствительностью к электрическим нолям переменного тока обладают обыкновенные скаты и акулы. Напряженность поля в десятые доли микровольта на 1 см оказалась достаточной, чтобы вызвать у акул рефлекс смыкания век (моргание глазами) и изменить у морской лисицы ритм дыхания. Выяснилось, что высокая чувствительность этих рыб к электрическим полям обусловлена наличием у них специфических рецепторов - так называемых ампул Лоренцини. При нарушении иннервации их чувствительность понижалась в несколько десятков раз. Ученые пришли к выводу, что ампулы Лоренцини являются у акул и скатов электрорецепторами.
В дальнейших экспериментах исследовалась чувствительность акул и скатов к электрическим полям естественного происхождения. Реакция рыб отмечалась с помощью электрокардиограммы, снимаемой с двух электродов, помещенных в перикард сердца. Если рыбы воспринимали электрические поля, мышечный потенциал сердца значительно изменялся - уменьшались его амплитуда и частота следования импульсов. Оказалось, что в благоприятных условиях акулы и скаты чувствительны к электрическим полям напряженностью в сотые доли микровольта на сантиметр, т. е. примерно в 10 раз более чувствительны к воздействию электрических полей, чем показывали опыты, проводившиеся другими способами.
При изучении ориентации слабоэлектрических рыб Лиссман установил, что некоторые из них воспринимают исключительно слабые электрические токи, возникающие в теле в результате индукции при движении в магнитном поле. Эта способность позволяет рыбам ориентироваться в таких полях. Гимнотуса, например, можно было приучить кормиться в участке аквариума, вблизи которого находился небольшой магнит. У гнатонемуса и гимнарха движение магнита вне аквариума на некотором расстоянии вызывало характерную двигательную реакцию.
Высокая чувствительность к электрическим полям позволяет слабоэлектрическим рыбам находить и различать в воде объекты, определять соленость воды, использовать разряды других рыб в межвидовых и внутривидовых отношениях. В основном мир ощущений у этих рыб электрический.
У разных видов рыб реакция на внешние электрические поля варьирует в очень широких пределах. Наибольшей чувствительностью обладают представители слабоэлектрических видов: у гимнарха она достигает 0,01 мкВ, стернарха - 0,02 и обыкновенных скатов - 1,2 мкВ на 1 см, а у некоторых сильноэлектрических рыб она значительно меньше: около 1 мВ на 1 см.
Чрезвычайно варьирует чувствительность неэлектрических видов. У некоторых сомовых и осетровых, имеющих специальные электрические рецепторы, она составляет десятки и сотни микровольт на 1 см, а у остальных неэлектрических рыб - десятки милливольт на 1 см.
Чувствительность отдельных участков тела рыб к электрическому току зависит от плотности расположения на них электрических рецепторов. У большинства рыб, имеющих электрические органы, например у гимнарха, такие рецепторы наиболее густо распределены на голове, спине и брюхе, у скатов - на брюхе. Для всех рыб характерно, что там, где находятся электрические органы, электрические рецепторы полностью отсутствуют.
Многочисленные исследования показали, что почти у всех слабоэлектрических и сильноэлектрических рыб специализированными электрорецепторами являются производные органов чувств боковой линии. Наиболее исследована электрорецепторная функция ампул Лоренцини. Этим образованиям присвоено имя итальянского ученого, описавшего их в 1678 г. Согласно его взглядам, эти ампулы - слизистые железы, находящиеся в коже рыб. Они относятся к системе органов чувств боковой линии и встречаются не только у хрящевых рыб (акул и скатов), но и у некоторых костистых рыб (морского тропического сома).
Ампулы Лоренцини (рис. 14) представляют собой довольно длинные (до нескольких сантиметров) трубочки-каналы, заполненные желеобразным веществом и заканчивающиеся на поверхности тела порами; внутри тела они образуют характерное расширение, в котором находятся чувствительные клетки. К каждой ампуле подходит несколько нервных веточек - обычно шесть, но иногда гораздо больше. У мраморного электрического ската их количество колеблется от 26 до 32.
Функциональное назначение ампул Лоренцини долгое время оставалось неясным. Их считали органами, выделяющими слизь, а в конце XIX в.- рецепторами гидростатического давления, предназначенными для ощущения глубины погружения рыбы. Это мнение подтверждалось отсутствием у скатов и акул плавательного пузыря - гидростатического органа костистых рыб. Правда, позднее выяснилось, что у морского тропического сома, обладающего плавательным пузырем, имеются и ампулы Лоренцини.
Электрофизиологические исследования ампул не внесли ясность в этот вопрос. Одни исследователи считали, что функция ампул терморецепторная, категорически отвергая их механорецепторную роль. Другие, наоборот, утверждали, что это механорецепторные органы, воспринимающие разницу давления внутри и снаружи ампул. Было также выдвинуто предположение об их хеморецепторной функции. Обнаружилась высокая чувствительность ампул к изменению концентрации солей в морской воде. Появилась гипотеза об электрической чувствительности этих органов.
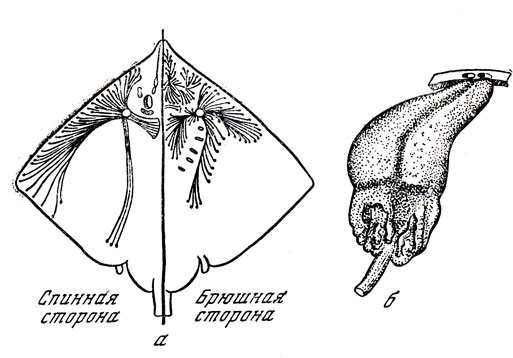
Рис. 14. Ампулы Лоренцини: а - расположение ампул Лоренцини на теле морской лисицы (точками на концах трубочек отмечены места, где ампулы открываются наружу); б - две ампулы Лоренцини у акулы (сверху отверстия на коже)
Однако только последующее изучение поведения рыб, в том числе эксперименты Дийкграфа и Кальмджина, о которых уже упоминалось выше, позволили выяснить электрорецепторное назначение ампул Лоренцини. Специальные электрофизиологические исследования подтвердили высокую чувствительность ампул к электрическим полям - приблизительно 0,1 мкВ на 1 см. Если один электрод (зонд) помещался в проток ампулы, а другой - на тело рыбы, то для "срабатывания" ампулы было достаточно действия электрического тока силой всего в 0,005 мА. Если же зонд смещали в сторону от поры ампулы всего на 0,5 мм, то для достижения такого же результата силу тока приходилось заметно увеличивать. Это свидетельствует о том, что электрический ток в основном протекает по трубочке ампулы.
Описываемое явление хорошо согласуется с морфологическими и биофизическими свойствами ампул Лоренцини. Установлено, что электрическое сопротивление стенок ампулы в 160 тыс. раз больше сопротивления желе, заполняющего ее. Электропроводность морской воды и желе в протоках ампулы приблизительно одинакова, остальных частей тела рыбы - примерно в 2 раза меньше. Доказано, что протоки ампул Лоренцини являются хорошо изолированными проводниками, по которым электрический ток передается с небольшими потерями. В связи со специфическим строением ампулы электрические разряды высокой частоты затухают в ее канале быстрее, чем разряды низкой частоты. Поэтому длинные ампулы Лоренцини могут воспринимать в основном низкочастотные разряды, а короткие - высокочастотные. Таким образом, ампулы Лоренцини представляют собой электрорецепторы. Однако они чувствительны и к другим раздражителям, прежде всего к механическим воздействиям. Возможно, что эти ампулы являются не только электрорецепторами, но и механорецепторами.
Как уже говорилось, высокой чувствительностью к электрическому току обладают, кроме акул и скатов, слабоэлектрические рыбы, например гнатонемус, гимнарх. У этих рыб имеются электрические рецепторы различных типов, образованные подобно ампулам Лоренцини, из органов чувств системы боковой линии. Наиболее чувствительны к электрическому раздражению ампулярные рецепторы: электрорецепторы гимнарха воспринимают напряженность электрического поля в 0,01 мкВ на 1 см.
Все исследованные электрические рецепторы слабоэлектрических рыб спонтанно генерируют импульсы определенной частоты - осцилляции. Они не зависят от разрядов электрических органов рыб и сохраняются даже в том случае, если кусочки кожи, на которых расположены рецепторы, изолированы. Однако по мере приближения к рецепторам объектов, создающих внешнее электрическое поле, частота осцилляций изменяется.
Функции электрических рецепторов слабоэлектрических рыб различны. Одни непрерывно работают как гальванометры (как бы замеряя величину тока); другие же воспринимают только изменение напряжения внешнего электрического поля, определяя его фазу, т. е. работают аналогично осциллографу.
Каким же образом осуществляется анализ информации, поступающей к рыбам извне и воспринимаемой их электрическими рецепторами? Известно, что органами боковой линии рыб управляют мозжечок и продолговатый мозг. Именно в них осуществляется анализ электрической информации. В связи с этим мозжечок у некоторых слабоэлектрических рыб исключительно сильно развит. Так, у мормируса относительная величина мозжечка больше, чем у любых других позвоночных животных. Он наиболее развит у сомовых рыб, которые, как и слабоэлектрические рыбы, обладают электрорецепторами. Степень развития "электрических" долей мозга слабоэлектрических и неэлектрических рыб хорошо видна на рис. 15.
У некоторых электрических и слабоэлектрических рыб, обладающих "электрическим мироощущением", относительно развиты боковые доли продолговатого мозга. Изучение этих отделов с точки зрения электрофизиологии позволило выявить несколько типов нейронов, по-разному реагирующих на внешнее электрическое поле, на движение в воде предметов с разной электропроводностью, а также различно адаптирующихся к этим раздражителям. Оказалось, что в зависимости от движения объектов активность одних нейронов электрического центра повышалась, а других, наоборот, понижалась. Благодаря этому рыба получала информацию о положении и перемещении в воде разнообразных объектов.
Для передачи любой информации от воспринимающих элементов к анализатору необходим соответствующий код - система условных обозначений. У человека это слова, отдельные фразы, термины, буквы, цифры, знаки и т. д. У рыб подобная система неизмеримо проще, но тем не менее нервные волокна, передающие электрическую информацию от воспринимающих рецепторов в нервные центры, "кодируют" ее разнообразными способами. Это отчасти можно объяснить тем, что электрорецепторы работают в комплексе с электрическими органами.
При одновременной работе электрических органов и электрорецепторов кодирование информации происходит довольно сложным путем. Так, представители гимнотовидных рыб в основном используют два типа кодирования: числовое" и "вероятностное". У этих рыб с низкочастной разрядной деятельностью (3-5 импульсов в секунду) нервные волокна как бы отвечают на каждый импульс цепочкой нервных импульсов. У электрического угря, например, эта цепочка состоит из 15 импульсов в секунду. Кодирование информации об изменении интенсивности внешнего электрического поля осуществляется путем изменения количества импульсов в цепочке - "числовое".
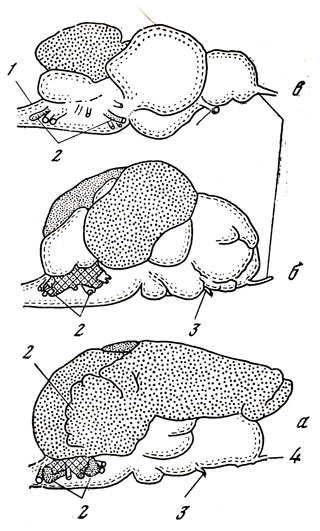
Рис. 15. 'Электрические' зоны мозга слабоэлектрических рыб (заштрихованные участки): а - мормирус, б - гимнарх, в - сом; 1 - позвоночный столб, 2 - нерв боковой линии, 3 - зрительный нерв, 4 - обонятельный нерв
У гимнотовидных рыб с высокой частотой разрядной деятельности электрических органов обнаружен другой способ кодирования - "вероятностный". При каждом электрическом импульсе импульсы в нервном волокне либо отсутствуют вообще, либо возникает один импульс (у некоторых рыб может возникнуть несколько). Кодирование электрической информации осуществляется в результате возникновения нервного импульса в зависимости от изменения интенсивности внешнего электрического поля.
Есть еще один способ кодирования - "условно-частотный". Нервные волокна проявляют спонтанную активность, не согласованную с разрядами электрического органа. Если во внешнее электрическое поле попадают какие-либо объекты, нервные волокна определенным образом изменяют частоту следования разрядов электрического органа.
В последнее время в СССР и за рубежом были проведены многочисленные исследования, посвященные кодированию электрической информации нервными волокнами. Интерес к этому вопросу закономерен. Зная механизм кодирования, можно понять, как протекает в организме анализ поступающей извне информации и каким образом раздражители различаются во времени и пространстве.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'