Глава 4. Донные, придонные и придонно-пелагические рыбы
Изучение рыб только по коллекционным образцам и так называемым «полевым биологическим анализам» (длина и масса тела, врелость гонад, наполнение желудков и т. п.) пикак по может считаться полноценным - оно почти не дает представления ни об их образе жизни, ни, тем более, поведении. Эти аспекты биологии особенно важно знать, когда имеешь дело с донно-придонными рыбами,- ведь они отличаются исключительным разнообразием взаимоотношений и поведенческих реакций. Если еще учесть, что рыб, поднятых с глубины, трудно содержать в аквариумах, становится понятным, почему об их поведении до сих пор известно так мало. Почитайте, к примеру, «Жизнь животных» — соответствующие разделы часто содержат лишь описания внешнего вида и сведения о глубине и месте поимки... Между тем в последние годы появилась прекрасная возможность преодолеть эти затруднения — она связана с использованием автономных обитаемых подводных аппаратов.
Такие аппараты есть и в Институте океанологии, и два из них (они рассчитаны на глубину до 2000 м) даже носят вполне ихтиологическое название «Pisces» (по-латински «рыбы»). К превеликому сожалению, ни я, ни кто-либо из коллег-ихтиологов на этих аппаратах-рыбах не погружался (в основном они обслуживают геологов), а наблюдения, сделанные нашими «штатными подводниками», надо прямо сказать, большой научной ценности не представляют (трудно поверить на слово, например, рассказу о том, что в Красном море не то винцигуерии, не то миктофиды зарываются в грунт на большой глубине). Конечно, фотографии и видеозаписи более информативны, но без осмысленного выбора нужного ракурса они далеко не всегда позволяют точно определить запечатленный объект.
В то же время другими исследователями уже выполнено немало ценных наблюдений над поведением глубоководных рыб. Приведу несколько примеров. Погружаясь в 1975 г. на аппарате «Эдвин» на материковом склоне северо-восточного побережья США, Дэн Коэн определил скорость антиморы, уходившей от преследования аппарата (39,2 см/с), и рассчитал, что расход энергии на плавание (3,6.105 эрг/с) у нее примерно одинаков с форелью того же размера - рыбой, известной своими скоростными качествами, В 1977 г. он же впервые сфотографировал па глубине около 2000 м самку речного угря, пришедшую на нерест к Багамским островам.Севастопольские ихтиологи Георгий Головапь и Николай Пахоруков в 1979 г. проследили с аппарата «ТИНРО», как на одной из гор подводного хребта Наска в Юго-Восточной Пацифике беспузырные окуни вместе с лангустами уходили с плоской вершины гайота на его склоны при подтоке с севера обедненной кислородом воды и снова появлялись па ней, когда ситуация менялась и на вершину вновь поступала вода, богатая кислородом.
...Я продолжаю надеяться, что и мне доведется когда-нибудь увидеть глубоководных рыб в естественной для них обстановке, А пока я наблюдал их в лучшем случае полуживыми - умирающими после изъятия из родной стихии.
Согласно принятому в этой книге определению «открытого океана» счет его донно-придонным биотопам начинается с материкового склона. Рассматривая субконтинентальный бентопелагический комплекс ихтиоценов, нужно иметь в виду, что в связи с сильной расчлененностью склона, препятствующей успешным тралениям, население мезо- и батибентали изучено значительно хуже, чем сообщества всех прочих крупномасштабных биотопов. Тем не менее изучение батиальных рыб в последние годы значительно продвинулось.
Прежде всего следует отметить, что в самых верхних горизонтах мезобентали настоящие среднеглубинные рыбы сосуществуют с сублиторальными, примесь которых в ихтиоцене может быть очень значительной. Эта переходная зона должна рассматриваться поэтому как экотон. Вблизи 200-метровой изобаты виды, общие для эпи- и мезобентали, настолько многочисленны, что нет смысла перечислять их здесь. Некоторые эврибатные формы встречаются в очень широком диапазоне глубин. Многие из них представлены у нижних пределов своего вертикального распространения более крупными особями, чем на мелководьях, для других, имеющих широкие ареалы, характерно увеличение глубины обитания в тропической зоне, но в целом мелководные и среднеглубинные виды обычно неплохо различаются по предпочитаемой глубине. Поэтому обыкновенного катрана (Squalus acanthias), например, наиболее обычного на глубине 20—200 м, относят к сублиторальным видам, а черного палтуса (Reinhardtius hippoglossoides), во взрослом состоянии концентрирующегося на 150—800 м, — к мезобентальным.
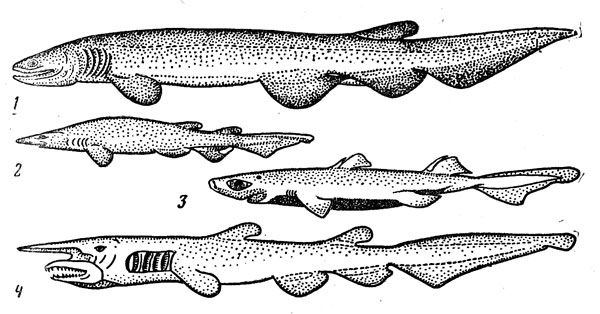
Рис. 31. Придонные акулы: плащеносная (1), глубоководная кошачья (2), этмоптерус (3), акула-домовой (4).
Мезобентопелагический комплекс ихтиоценов исключительно разнообразен по составу. Среди характерных для него щележаберных рыб нужно отметить плащеносных (сем. Chlamydoselachidae), колючих (Squalidae) и кошачьих (Scyliorhinidae) акул, а также ромбовых скатов (Rajidae).
Очень своеобразная плащеносная акула (Chlamydoselachus anguineus) (рис. 31) — представитель одной из наиболее примитивных групп щележаберных — имеет угревидно удлиненное тело и жаберные щели, прикрытые кожистыми складками (их по шесть с каждой стороны вместо пяти у большинства акул). Она может достигать 2 м в длину, но половозрелость наступает при значительно меньших размерах — у самцов около 1 м, самок — 1,3 м, Размножается путем яйцеживорождения, принося от 8 до 12 детенышей длиной 40 см, причем считается, что вынашивание эмбрионов продолжается очень долго (год или даже два). Эта редкая акула обитает на материковом склоне и подводных поднятиях (глубина ловов варьирует от 120 до 1300 м) в умеренно теплых и субтропических Районах всех океанов. Близкие к ней и тоже не очень часто встречающиеся семи- и шестижаберные акулы (рода Heptranchias и Hexanchus) также достигают довольно крупных размеров (1,5—4,8 м) и ведут придонный образ жизни в широком диапазоне глубин шельфа и склона.
Колючие акулы, напротив, принадлежат к числу самых обычных обитателей мезобентопелагиали — в ее пределах встречаются 58 из 68 видов семейства (еще 10, как уже говорилось, живут в пелагиали без связи с дном), причем 46 видов обитают только в этом биотопе, а три являются общими для мезо- и батибентопелагического комплекса Один из них встречается и над ложем океана — португальской акуле Centroscymnus coelolepis, поимка которой зарегистрирована в 3675 м от поверхности, принадлежит рекордное по глубине нахождение среди всех хрящевых рыб (правда, еще глубже — па глубине более 6000 м наблюдали из батискафа не определенных до вида колючих акул, отнесенных к роду Centrophorus). Совокупный ареал семейства простирается от Арктики до Субантарктики. Очень широко распространены и многие виды, обнаруженные во всех трех океанах: эврибатный обыкновенвый катран, например, обитает в умеренных водах обоих полушарий, а в тропиках его почти повсюду замещает малошипый катран (Squalus mitsukurii).
Колючие акулы сильно различаются по размерам. Длина тела у мелких видов рода Etmopterus не превышает 25—30 см (к этому роду относятся самые мелкие из ныне живущих акул: Е. carteri и Е. perryi становятся половозрелыми при длине 16—18 см и вырастают всего лишь да 21 см), а у гигантских представителей рода Somniosia достигает 6 м и более. Наибольшая из достоверно измерена ных полярных акул (S. microcephalus) имела 6,4 м в длину при массе около 1 т, но есть сообщения и о еще более крупных экземплярах (К. Г. Константинов отмечал поимку в 1956 г. двух 8-метровых особей у берегов Мурмана) Этот вид распространен в северной Атлантике (на юг до залива Мэн и Пиренейского полуострова), а также у Юна ной Африки и субантарктических островов. В высоких широтах полярная акула нередко (особенно зимой) встречается на мелководьях — у Гренландии эскимосы промышляют ее через проруби в припае, а в умеренно теплых водах предпочитает глубины склона (до 1200м). Она питается разнообразной рыбой, часто нападает на тюленей может пожирать любые отбросы и падаль. Долгое время считалось доказанным, что эта акула в отличие от вся других представителей семейства размножается, откладывая в воду мягкие яйца (это представление зафиксирован во всех учебниках ихтиологии), но в действительности oна яйцеживородяща, как и все ее родичи: из самки длиной около 5 м было извлечено 10 готовых к рождению эмбрионов длиной 37 см.
Некоторые колючие акулы (в том числе обыкновенные катран) ведут стайный образ жизни и совершают значительные по протяженности сезонные миграции. Интересно, что собравшиеся в стаю особи одного из самых мелких видов — карибско-мексиканского Etmopterus virens, живущего на глубине 350—465 м, могут совместно атаковать крупную добычу, в частности больших кальмаров, и разрывать ее на куски. Эти акулы, подобно многим другим сквалидам, имеют фотофоры и обладают способностью к свечению. Рост колючих акул практически не изучен. Данные по возрасту имеются только для обыкновенного катрана, который достигает зрелости на 10—20-м году и живет по меньшей мере до 25—30 лет. Плодовитость у разных видов варьирует от 2 до 20 эмбрионов.
Большое семейство кошачьих акул, включающее около 90 видов, также очень характерно для субконтипентальной мезобентали, в которой они представлены 62 видами (остальные обитают в прибрежной зоне). На значительных глубинах особенно обычны представители рода Apristurus, из которых четыре даже встречены глубже 1500 м (A. microps добывался только в диапазоне глубин от 1000 до 2000 м). Все кошачьи акулы ведут оседлый образ жизни и не совершают больших перемещений, а их apeaлы, как правило, невелики по площади. Они имеют небольшие размеры: многие виды достигают половой зрелости уже при длине 20 см, а самые крупные из среднеглубинных форм — проникающие с шельфа на склон акулы-кошки рода Scyliorhinus — не превышают 1,2 м в длину.
В этом семействе наблюдается значительное разнообразие типов размножения. Наиболее примитивный способ состоит в последовательной откладке на субстрат каждого оплодотворенного яйца, заключенного в прочную капсулу, которая прикрепляется ко дну роговыми нитями; развитие зародыша продолжается без всякой охраны в течение продолжительного времени — около года. У других видов яйца упаковываются в капсулы по нескольку штук и про ходят начальные этапы развития в яйцеводах матери, так что время между откладкой капсулы на дно и вылуплением детенышей сокращается до нескольких недель. Есть реди кошачьих акул и яйцеживородящие виды, у которых эмбрионы остаются в яйцеводах вплоть до полного рассеивания желточного мешка и рождаются на свет вполне товыми к самостоятельному существованию. Примеры всех трех способов размножения можно найти, в частности У разных видов из родов Halaelurus или Galeus. Так, энский галеус (G. nipponensis) откладывает одиночные яйца, черноротая акула (G, melastomus), обычная на материковом склоне Европы, вынашивает в яйцеводах до 13 яиц, упакованных в капсулы для последующей откладки на дно, а западноафриканский вид G. polli способен в яйцеживорождепию. Мало того, Ф. Ф. Литвинов сообщил мне, что он наблюдал у последнего вида даже внутриутробное поедание неоплодотворенных яиц, которое считалось до сих пор свойственным только ламноидным акулам (об этом уже говорилось в гл. II). В эмбриональной оофагии и каннибализме подозревалась, правда, и ложнокунья акуле (Pseudotriakis microdon) — довольно крупный (длина до 3 м) мезобентальный вид, известный с глубины 200-1500 м во всех океанах. Эта акула приносит всего 2—4, но весьма солидных по размерам (70—85 см) детенышей, наряду с которыми в яйцеводах находили большие запасы желтковых яиц (до 20 тыс. в каждом яйцеводе у самки длиной 2,8 м); это подозрение остается, однако, недоказанным, так как желтка в желудках эмбрионов пока не обнаружено.
Из других глубоководных акул нужно прежде всего упомянуть митсукурину, или акулу-домового (Mitsukurinai owstoni), выделяющуюся своей необычной, прямо-таки неправдоподобной внешностью: ее сильно удлиненное рыло имеет форму заостренного сошника, а челюсти обладают способностью выдвигаться далеко вперед, что связано по-видимому, с питанием донными животными. Эта акула известная по считанным поимкам, достигает 4 м в длину. Она ловилась на глубине до 550 м, но обломанный зуб, несомненно, принадлежащий этому виду, был извлечен однажды из пробитого телеграфного кабеля, проложенного по дну в 1350 м от поверхности. Пара митсукурин была доставлена в Атлантический институт рыбного хозяйства и океанографии в Калининграде, и чучело одной из них украшает сейчас музей этого учреждения.
Отдельные виды, входящие в глубоководные бентопелагические комплексы, есть и в некоторых преимущественно шельфовых семействах акул. Среди куньих акул (сем. Triakidae), например, один вид (Jago omanensis) известен по левам на глубине 110—2195 м. Только в водам свала глубин (600—1000 м) обитает большеглазая песчаная акула Odontaspis noronhai. Придонный вид, тяготеющий к глубинам, есть даже среди серых акул: большеносая акула (Carcharhinus altimus), встреченная во всех океанах придерживается главным образом переходной зоны между эпи- и мезобенталью (обычна на глубине 90—430 м). По внешним признакам она очень близка к одному из прибрежных видов (С. plumbeus), от которого отличается лишь мелкими деталями строения.
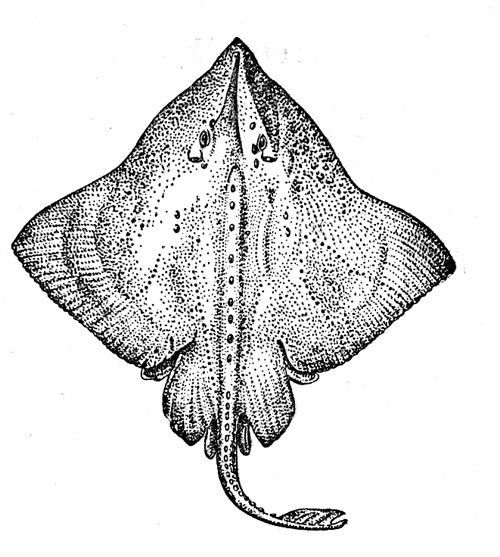
Рис. 32. Северный ромбовый скат.
Ромбовые скаты (рисунок 32) — типично донные рыбы, Они проводят свою жизнь, лежа на грунте, в который могут закапываться так, что на поверхности остаются только глаза, брызгальца и часть спины, или плавая в придонном слое. Их пищу составляют различные рыбы и беспозвоночные (преимущественно ракообразные), причем при охоте на плавающую добычу скаты наплывают на жертву сверху и прижимают ее ко дну своим диском, а потом уже заглатывают. В этом семействе, систематику которого интенсивно разрабатывают сейчас специалисты разных стран — М. Штеманн из Гамбурга, владивостокский ихтиолог В. Н. Долганов, японец X. Исихара и др., еще недавно числили 170—180 видов, а теперь их насчитывают уже более 200 (почти половина всего подотряда скатов), однако и эта цифра будет, без сомнения, увеличена. Ромбовые скаты живут во всех океанах и морях — их ареал простирается от Арктики до Антарктики. Они особенно характерны для холодных и умеренно теплых районов, но встречаются и в тропической зоне, где обитают обычно на значительной глубине. Большая часть видов (в том числе почти все представители самого обширного рода Raja, включающего около 140 видов) имеет центром вертикального диапазона распределения зону, расположенную на границе шельфа и склона, и относится, таким образом, к эпимезобентальной группировке. Наиболее глубоководные формы, которые принадлежат большей частью к родам Bathyraja (это крупные скаты длиной около 2 м) и Breviraja (их длина 30—40 см), вполне обычны и в батибентали: на рекордной глубине 2400—2950 м выловлены в Бискайском заливе типовые экземпляры Bathyraja pallida. Ромбовые скаты размножаются, откладывая на дно яйца, заключенные по одному в роговые капсулы с характерными трубчатыми отростками по углам. Как показали наблюдения в аквариуме, самка обычно выметывает яйца парами с перерывом в 1—5 дней, и общий период размножения может быть очень длительным. Число отложенных капсул, по-видимому, сильно варьирует у разных видов: от 4—5 до 50 и более. Инкубация яиц длится от 4 до 14 месяцев. Скаты принадлежат к долгоживущим видам. По данным В. Н. Долганова, батиальные виды, обитающие в дальневосточных морях, достигают половой зрелости в возрасте 3—10 лет, а живут от 7 до 18 лет (в зависимости от размера вида).
Вне шельфов представлены и некоторые другие группы скатов, в том числе три тропических семейства, близкородственные ромбовым скатам — Anacanthobatidae (нитерылые скаты), Arhynchobatidae (безрылые) и Pseudorajidae (ложноромбовые), которые в полном составе принадлежат к мезобентальиому ихтиоцену. К нему же относятся очень своеобразные шестижаберные скаты (сем. Нехаtrygonidae) — хвостоколы с шестью жаберными щелями. Первый их представитель — Hexatrygon bickelli — был найден в 1981 г. выброшенным на берег у Порт-Элизабет (Южная Африка); в том же году другой вид этого рода был описан по 2 экз. из Южно-Китайского моря, а затем к ним добавилась пара рыб, пойманных у островов Рюкю и Гавайских в диапазоне глубин от 350 до 1000 м. Еще один молодой самец, выловленный в Индийском океане вблизи острова Ява, был обнаружен недавно Ю. Н. Щербачевым среди не определенных рыб в коллекции Зоологического музея МГУ.
На довольно больших глубинах обитают и некоторые электрические скаты из семейства гнюсовых (Torpedinidae), обладающие, как хорошо известно, своеобразными электрическими органами, расположенными по бокам тела между головой и грудными плавниками. Так, многие виды рода Torpedo, обычные на шельфе, могут встречаться вплоть до глубины 400—500 м, а Т. tokionis из вод Японии и индийский Benthobatis moresbyi (сем. Narkidae) живут только на склоне (220—1100 м). Как это ни странно, некоторые гнюсы (например, Т. tremens из юго-восточной Пацифики) способны и к пелагическому образу жизни; этот факт вызывает удивление, поскольку все электрические скаты плохие пловцы: их единственным движителем служит короткий хвостовой стебель с небольшой лопастью хвостового плавника.
Когда мне впервые довелось увидеть живого гпюса в траловом улове (это было на горе Большая - одном из гайотов подводного хребта Наска), я недолго боролся с искушением испытать на себе действие его электрического разряда. Особого геройства в этом, конечно, не было - ведь во всех сводках и руководствах вполне четко сказано, что максимум напряжения «батарей» этих рыб не превышает бытовых 220 вольт (я и сам писал об этом в «Жизни животных»). Тем не менее я взял ската в руки внутренне готовый к тому, что сейчас ощутимо «дернет», но... Гнюс лишь вяло шевельнул хвостом, явно не желая показывать свое умение. Я помял ему брюхо и спину, пощекотал пальцем голову. Никакого эффекта. Отложив ската в сторону, я стал выбирать из улова, надо сказать довольно скудного, других рыб, заслуживавших внимания, и этот отбор занял минут десять. Собравшись в лабораторию, я взял ведро с рыбой и небрежно ухватил гнюса за хвост, на что он тут же ответил довольно чувствительным (жаль, не было вольтметра) и совершенно неожиданным разрядом, на который я отреагировал не менее быстро, но, к сожалению, отнюдь не лучшим и, главное, столь же неожиданным для самого себя образом — одним движением выбросил обидчика за борт.
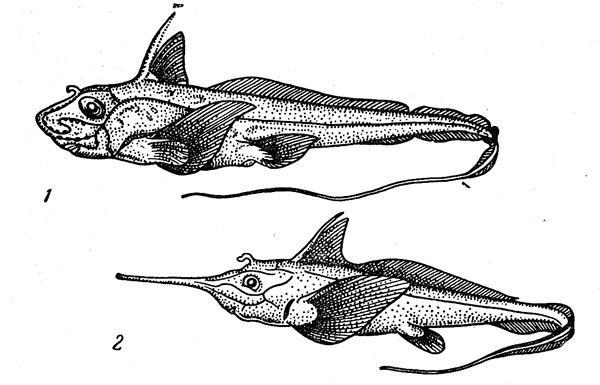
Рис. 33. Европейская химера (1) и ринохимера (2).
Дать правдоподобное объяснение происшедшему не так уж трудно: по-видимому, электрический скат еще в трале израсходовал всю емкость своих батарей и на подзарядку севших элементов потребовалось время, которого как раз хватило до моего вторичного прикосновения.
Как следует из сказанного выше, доминирующие в мезобентопелагических комплексах группы щележаберных рыб существенно различаются по тесноте связи с твердым субстратом — ромбовые скаты, проводящие большую часть времени непосредственно на дне, могут быть отнесены (с очень небольшой натяжкой) к бентосу, кошачьи акулы, также нередко покоящиеся на грунте, — к нектобентосу, а колючие акулы — к бентонектону. Соответственно этому колючие акулы значительно чаще встречаются в пелагиали (некоторые из них даже над большими глубинами), чем представители двух других семейств. Пространственное распределение этих рыб отражает различия их экологии: переход от донного образа жизни к придонно-пелагическому сопровождается изменением типа размножения (от откладки капсулированных яиц к яйцеживорождению) и состава пищи (от потребления бентических беспозвоночных к охоте на рыб и кальмаров в водной толще).
Еще одна очень характерная группа глубоководных донных рыб — цельноголовые, которые представлены в мезо- и батибентали двумя семействами (рис. 33). Химеровые (Ghimaeridae) обитают преимущественно в верхней части материкового склона на глубине от 200 до 700— 1000 м. Изученная лучше других европейская химера (Chimaera monstrosa), обитающая в восточной Атлантике (один экземпляр был пойман в 1957 г. в советских водах — у берегов восточного Мурмана), достигает 1,5 м в длину. Другие представители семейства (в нем числятся два рода и более 20 видов), как правило, также имеют довольно ограниченное распространение, хотя общий его ареал охватывает батиаль всех тропических и умеренно теплых районов Мирового океана, причем у краев этой широтной зоны химеры встречаются и на относительно небольших глубинах. Будучи медлительными и неповоротливыми, эти рыбы постоянно держатся у дпа. Они плохие пловцы, передвигающиеся только используя в качестве весел свои вееровидные грудные плавники. Пищу химер составляют донные беспозвоночные и рыбы, а размножаются они, откладывая на дно веретеновидные яйцевые капсулы, которые прикрепляются к субстрату специальными роговыми нитями.
Близкое семейство носатых химер (Rhinocnimaeridae), имеющих очень длинное заостренное рыло, также специфично для материкового склона. Наиболее глубоководными среди 6 видов этой группы являются гарриотты (род Harriotta), опускающиеся до глубины 2600 м.
Перейдем теперь к костным рыбам. Совершенно особое место среди них занимает латимерия, или целакант (Latimeria chalumnae), — единственный современный представитель большой группы кистеперых рыб, когда-то — с нижнего девона (400 млн лет назад) до верхнего мела (65 млн лет назад) — процветавших в морских и пресных водоемах Земли, а затем полностью исчезнувших из палеонтологической летописи и считавшихся давным-давно вымершими. Именно поэтому поимка живого целаканта у берегов Южной Африки в 1938 г. стала одной из крупнейших биологических сенсаций нынешнего столетия, впрочем, эта история слишком хорошо известна, чтобы еще раз пересказывать ее здесь (с ней можно ознакомиться, прочитав книгу проф. Дж. Смита «Старипа — четвероног»).
Сейчас считается установленным, что латимерии, как это ни удивительно, нормально обитают только в одном и к тому же очень ограниченном участке Мирового океана — у бесшельфовых вулканических островов Гранд-Комор и Анжуан (из группы Коморских островов), расположенных в северной части Мозамбикского пролива, а первая из пойманных рыб лишь случайно заплыла так далеко на юг. К настоящему времени в руки ученых попало уже около сотни латимерии, и все они были пойманы коморскими рыбаками на традиционные примитивные удочки, предназначенные в основном для лова руветты (жир этой рыбы используется ими для изготовления противомалярийной мази для растирания). Промысел ведут ночью с небольших лодок (типа каноэ с аутриггером), не удаляющихся от берега больше чем на километр. Этого достаточно, чтобы занять место над свалом глубин и опустить крючок, наживленный куском прометеевой рыбы, в придонный слой. Именно там, среди скал на глубине от 70 до 600 м, ловятся целаканты (местные жители называют их «гомбесса джомоле»), но чаще всего они берут приманку на глубине 150—300 м. Есть предположение, что Дневные убежища латимерии приурочены к подводным пещерам, в которые по трещинам просачивается пресная вода с гор. Пищу целакантов составляют рыбы (бентопелагические светящиеся анчоусы, полимиксии и др.) и каракатицы. Обычно добывают крупных латимерии длиной 90—200 см и массой 10—95 кг (предполагается, что их максимальный возраст составляет около 10 лет), однако один Раз была добыта и совсем небольшая, по-видимому, годовалая рыба, имевшая всего 42 см в длину при массе 800 г, размножение происходит путем яйцеживорождения (как у многих акул) — в яичнике одной самки были обнаружены 5 почти готовых к рождению эмбрионов длиной 30-33 см с еще не втянувшимися желточными мешками дна метром 8—13 см.
Все остальные виды, входящие в состав океанических донно-придонных ихтиоцепов, относятся к костистым рыбам. Рассмотрим их в порядке системы, принимаемой сейчас большинством ихтиологов.
Отряд спиношипообразных полностью ограничен в своем распространении мезобатибенталыо. Нотакантовые (сем. Notacanthidae) встречаются во всех океанах в довольно широком диапазоне глубин (от 125 до 3700 м), но большинство видов тяготеет все-таки к верхней части склона (200—1300 м). Они держатся в узком придонном слое (как показали подводные наблюдения, эти рыбы плавают обычно в наклонном положении, так что голова находится у самого грунта) и питаются бентосными животными. Нотаканты, или спиношипы (род Notacanthus) (рис. 34), особенно часто поедают мшанок, гидроидных полипов, губок, актиний, голотурий, а многошипы (род Ро-lyacanthonotus) — ракообразных и полихет. Известно, что у многих видов самки численно преобладают над самцами и имеют более крупные размеры, п это дает основания для довольно правдоподобного предположения о возможном протандрическом гермафродитизме этих рыб. Их личинки — лептоцефалы имеют листовидно уплощенное удлиненное и прозрачное тело и достигают очень крупных размеров (до 1,8 м). При метаморфозе длина, по-видимому, сильно уменьшается, так как взрослые экземпляры большинства видов не превышают 50—60 см в длину (только Notacanthus chemnitzi может вырастать до 120 см).
Семейство галозавровых (Halosauridae), представители которого вылавливались на глубине 400—5000 м (обычно не глубже 3000 м, в абиссобенталь проникает только один вид рода Aldrovandia), мало отличается от нотакантовых по образу жизни. Их пищу также составляет бентос, но не седентарные (сидячие) формы, а главным образом ракообразные и полихеты. Одни виды предпочитают твердые грунты, другие — илистые участки. Особи некоторых видов, возможно, держатся стайками. Семейство насчитывает 16 видов, среди которых есть как циркумтропические формы (Halosauropsis macrochir, Aldrovandia affinis), более обычные в батибентали, так и узкораспространенные (многие виды рода Halosaurus), которые тяготеют к мезо-бентальным глубинам.
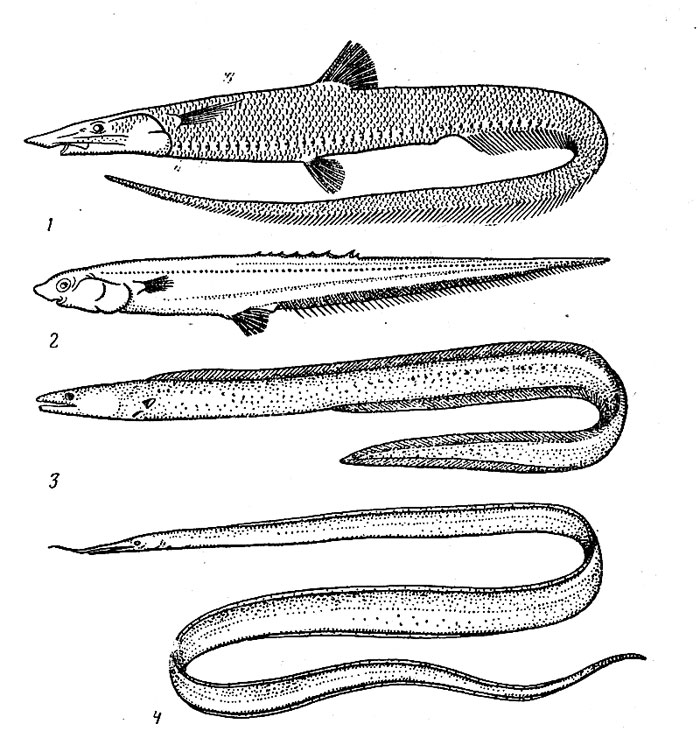
Рис. 34. Альдровандия (J), спиношип (2), слитножаберный угорь (3) и угорь венсфика (4).
Довольно обычны, а местами и многочисленны в батиальной зоне (преимущественно в пределах тропической области) донно-придонные угреобразные рыбы, принадлежащие к пяти семействам (всего в отряде их насчитывается 16) и различающиеся по глубине обитания. Только в верхней мезопелагиали — до глубины 300—400 м встречаются отдельные представители преимущественно мелководных острохвостых угрей (сем. Ophichthidae), в том числе Ophisurus serpens — наиболее крупный вид семейства, достигающий 2,5 м в длину, а также ложномуреновых угрей (Xenocongridae). Глубже 500—600 м редко опускаются и конгеровые угри (Congridae) — семейство, которое насчитывает около 30 родов и 100 видов, в большинстве своем шельфовых. Наиболее глубоководный из них — Coloconger scholesi, обитающий в западной части Тихого и в Индийском океанах, живет, однако, па глубине 600-1200 м.
Специфичны для мезобатибентали всего два семейства угрей — неттастомовые (Nettastomatidae) и слитножабер-ные (Synaphobranchidae). К первому из них принадлежат более 35 видов, из которых 10 известны сейчас только по личинкам — лептоцефалам. Взрослые рыбы достигают 1 м в длину. Все они имеют сильно вытянутое рыло, у некоторых оканчивающееся мясистым щупальцевидным отростком. Эти угри живут на глубине от 500 до 2000 м, причем чернохвостая неттастома (Nettastoma melanurum), а возможно, и другие виды устраивают норы в мягком грунте. Основной их пищей являются макропланктонные ракообразные.
Слитножаберные угри обычно обитают в том же батиметрическом диапазоне, хотя отдельные особи ловились и глубже 4000 м. Среди них есть как эпимезобентальные (виды рода Dissoma), так и чисто батибеитальные (Histiobranchus bathybius) формы; некоторые достигают крупных размеров (рекордная длина 1,8 м отмечена у Diastobranchus capensis из вод Южной Африки и Австралии). Это — хищники и пожиратели падали, питающиеся рыбой, головоногими моллюсками и ракообразными. Один из видов — обезьяний угорь (Simenchelys parasiticus) наряду с потреблением мелких амфипод и копепод может, как предполагают, нападать на крупных рыб и, прогрызая стенку тела, выедать их внутренности (возможно, впрочем, что объектом «нападения» были в действительности уже погибшие рыбы). Некоторые из слитножаберных угрей встречаются далеко за границами тропической зоны — вплоть до Исландии и Берингова моря, но эти районы могут, вероятно, считаться областью их стерильного выселения.
Исключительно своеобразные одночелюстные угри (сем. Monognathidae), у которых в отличие от всех прочих рыб верхняя челюсть полностью редуцирована, так что формально их можно назвать челюстноротыми с большой натяжкой, относятся тем не менее всего лишь к особому семейству, близкому к мешкоротам. В ихтиологических коллекциях разных стран сейчас насчитывается около 75 экз. моногнатов, и все они были исследованы датскими учеными И. Нильсеном и Э. Бертельсеном, получившими исключительно интересные результаты. Почти все рыбы длиной от 4 до 15 см были пойманы в толще воды на глубине от 200 до 2000 м, но более частая встречаемость в придонных горизонтах позволяет предположительно относить ату группу к бентопелагическому ихтиоцепу, тем более что самая крупная особь длиной около 30 см была поймана у дна (этот экземпляр, отличающийся от прочих черной окраской и хранящийся ныне в Зоологическом институте АН СССР, выловлен в районе Курильских островов). Семейство включает 14 видов, многие из которых известны по единичным особям. Единственную пищу мопогнатов составляют крупные креветки, которых эти рыбы, вовсе не способные к быстрому плаванию и к тому же лишенные органов зрения, обоняния (обонятельные розетки развиваются только у половозрелых самцов) и сейсмосенсориой системы, конечно, не могут ловить активным путем. Предполагается, что они подманивают добычу, выделяя в воду пахучий секрет специальной железы, расположенной па голове, и убивают близко подплывшую креветку, проткнув ее панцирь особым ростральным зубом, направленным вниз. Этот непарный зуб имеет трубчатое строение (внутренний канал открывается вблизи его острой вершины), и у его основания расположена железа, вырабатывающая ядовитый секрет. Подобный аппарат, гомологичный зубам ядовитых змей, является совершенно уникальным среди рыб и рыбообразных.
Отряд лососеобразных представлен па глубинах несколькими группами. Семейство серебрянковых (Argentinidae), содержащее около 35 видов, включает как бенто-пелагические, так и настоящие пелагические формы, живущие на средних глубинах. К водам, омывающим верхнюю часть материкового склона (на глубине 150—700 м, редко глубже), приурочены два рода — Argentina и Glossanodon. Это некрупные рыбы длиной 15—20 см, достигающие кое-где высокой численности. Почти все серебрянки днем держатся у самого дна, а ночью стаями поднимаются в толщу воды. Лучше других изучена большая аргентина (A. silus), обитающая в умеренных водах Северной Атлантики. Она питается зоопланктоном (эуфаузииды, мелкие креветки и др.), имеет большую продолжительность жизни и достигает максимального размера (60 см) к 25 годам. Половозрелой становится в возрасте 10—12 лет, нерестится в апреле — июне, выметывая 3500—7500 крупных пелагических икринок. Второй род семейства — глоссанодоны, или малозубые серебрянки, тяготеет к субтропическим водам.
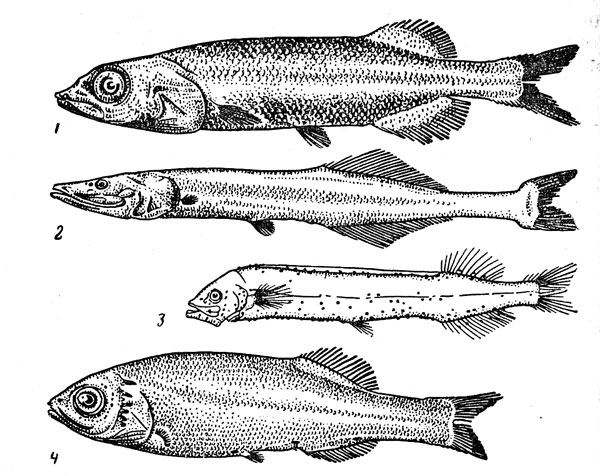
Рис. 35. Гладкоголововидные рыбы: гладкоголов (1), батиприон (2), фотостил (3), сирсия (4).
Гладкоголововидные рыбы (рис. 35) — группа, относящаяся к числу самых характерных для материкового склона и склонов подводных поднятий. В последние годы их изучение значительно продвинулось прежде всего благодаря работам Ю. И. Сазонова и Д. Маркла, но это относится главным образом к систематике, так как сведений об экологии отдельных видов до сих пор недостает. К семейству гладкоголовых (Alepocephalidae), не представленному только в Арктике и Антарктике, принадлежит около 90 видов (наиболее разнообразна фауна тропической Индо-Вестпацифики). Все они входят в состав бентопелагических ихтиоценов, однако их связь с дном выражена в разной степени. Одни виды, составляющие преобладающее большинство (представители родов Alepocephalus, Conoсага и др.), приурочены к узкому придонному слою, другие, например Photostylus pycnopterus, обитают и в значительном удалении от склона, третьи (Microphotolepis) даже совершают суточные вертикальные миграции, поднимаясь ночью к границе эпипелагиали. В целом диапазон батиметрического распространения семейства очень широк — от 100—150 до 5000 м, но наиболее обычны гладкоголовы на глубине 700—3000 м. В семействе преобладают виды, тяготеющие к нижней батиали, а число верхнебатиальных значительно больше, чем абиссальных (к последним относятся, по-видимому, роды Bellocia и Rinoctes), размеры гладкоголовов варьируют от 15—20 см до 1 м. В их питании преобладают планктонные объекты (очень обычны, в частности, «студенистые» формы — медузы, гребневики, пиросомы, а также ракообразные), но нередко поедается и бентос (моллюски, полихеты, иглокожие). Рост медленный — южный гладкоголов (Alepocephalus australis), например, достигает длины 70 см в возрасте 20 лет. Плодовитость колеблется от 20 икринок (у Rinoctes nasu-tus) до 10 тыс. икринок. Они имеют очень большой диаметр (2—7 мм) и, по всей вероятности, откладываются на дно.
Уникальной особенностью семейства платитроктовых (Platytroctidae), по внешнему виду очень сходных с гладкоголовами, служит наличие подкожного плечевого органа, продуцирующего светящуюся слизь. Этот орган открывается наружу маленькой кожной трубочкой, через которую слизь, содержащая люциферин и люциферазу, извергается наружу и распыляется в воде, где создает яркое светящееся облачко голубовато-зеленого цвета. У некоторых видов семейства есть и фотофоры, но не округлой формы, как у большинства других светящихся рыб, а в виде полосок-«шевронов», пересекающих, как правило, нижнюю поверхность тела. В семействе насчитывается около 40 видов, из которых большая часть описана в последние 20—25 лет. Платитроктовые — некрупные рыбы длиной от 12 до 30 см (наибольших размеров достигают представители рода Holtbyrnia). В своем преобладающем большинстве они обитают в бентопелагиали на глубине 700—2000 м, и лишь один вид — Platytroctegen mirus, распространенный в северной части Индийского океана, возможно относится к числу мезопелагических. Биология этих рыб практически не изучена.
Немаловажная роль в бентопелагических ихтиоценах принадлежит и некоторым макропланктонным рыбам, входящим в состав таких семейств, в которых преобладают настоящие пелагические виды, не имеющие никакой связи с дном. В отряде стомиеобразных такие виды есть среди гоностомовых (например, Triplophos hemingi и Gouostoma denndatum), топориковых (все 25 видов рода Роlyipnus, мавролик Maurolicus muelleri) (рис. 36), фотихто-вых (виды рода Polymetme), астронестовых (группа Astronesthes lucifer), меланостомиевых (два вида рода Odontostomias). Особое место в этом списке занимает мавролик (по всей вероятности, под приведенным выше названием объединяют 5—6 близких видов), который является одной из самых массовых рыб во многих участках материкового склона (в водах Норвегии он живет даже в глубоких фиордах) и у некоторых подводных поднятий. Наибольшая длина мавролика, широко распространенного над материковыми склонами и подводными поднятиями во всех океанах, достигает 6—6,5 см. Это стайная рыба, держащаяся днем в придонных слоях на глубине 200—600 м, но отрывающаяся от дна ночью и плотными косяками поднимающаяся в верхний 100-метровый слой. Мавролик созревает в годовалом возрасте и выметывает 200—500 довольно крупных икринок, причем размножение в умеренных водах обоих полушарий происходит в весенне-летнее время, а в тропиках идет круглогодично. Возраст этой рыбы не превышает 2—3 лет. Питается она копеподами и эвфаузиидами, а сама служит пищей многим хищникам.
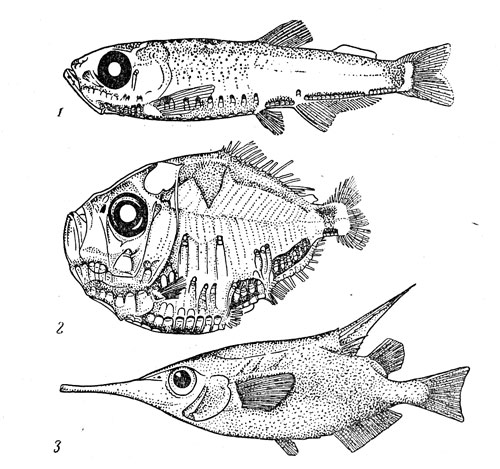
Рис. 36. Мавролик (1), топорик полипнус (3) и морской бекас (3).
К придонно-пелагическому комплексу средних глубин относятся и многие светящиеся анчоусы. Наиболее известен среди них лампаниктод (Lampanyctodes hectoris), обитающий в водах материкового склона Южной Африки, Австралии, Новой Зеландии и Чили и образующий в некоторых районах значительные скопления, которые используются промыслом. В темное время суток эта небольшая миктофида (длина до 6—7 см) совершает миграции в верхние слои воды и даже появляется у поверхности. Аналогичный образ жизни ведет Diaphus suborbitalis (наибольшая длина 8 см), распространенный у берегов Восточной Африки, Индонезии и Японии (он особенно многочислен в заливах Сагами и Суруга), а также над многими подводными поднятиями в Индийском и Тихом океанах. Нужно отметить, что во время ночного подъема диафус удаляется на 200—300 м от дна, иногда пересекая разнонаправленные течения, и остается тем не менее над своим поднятием, ко дну которого он вновь опускается утром. Как видно, эта рыба, во-первых, может ориентироваться в пространстве, не имея зрительного контакта с неподвижным субстратом, и, во-вторых, способна противостоять течениям, имеющим скорость порядка 1—2 узлов (т. е. 1,8—3,6 км/ч), чтобы сохранять свое положение относительно горы. Способ ориентации остается пока неизвестным. Во всех остальных отношениях этот вид не отличается от мигрирующих мезопелагических миктофид: он питается зоопланктоном и имеет короткий жизненный цикл — созревает к концу первого года жизни, а затем нерестится, выметывая порцию за порцией икру в течение нескольких месяцев вплоть до естественной гибели. Из других бентопелагических светящихся анчоусов, совершающих суточные миграции, следует упомянуть такие виды, как D. taaningi и Benthosema fibulatum, достигающие высокой численности (первый — над материковым склоном обоих берегов Атлантики, второй — в Индийском и Тихом океанах). Среди миктофид есть и виды, постоянно приуроченные к очень узкому придонному слою и не покидающие его даже ночью. Эти виды, например Diaphus adenomus и D. coeruleus, ловятся только донными тралами.
Придонные виды известны также в таких преимущественно пелагических группах, как семейство меламфаевых (Melamphaes acanthomus обитает только над склоном западного побережья Америки от Калифорнии до 20° ю. ш.) и подотряд глубоководных удильщиков, в котором все 4 вида семейства дицерациевых (Diceratiidae) приурочены к мезобентопелагиали, а представители рода Thaumatichthys (сем. Thaumatichthyidae) вообще ведут донный образ жизни. Тауматихт (Т. axeli), известный с глубины до 3600 м, поджидает свою добычу, которую он привлекает светящейся приманкой, расположенно внутри ротовой полости, лежа на дне, и заглатывает ей не предпринимая практически никаких усилий.

Рис. 37. Глубоководные аулопообразныс рыбы: ипнопс (1), батитифлопс (2) батиптер (3), бентозавр (4).
Отряд аулопообразных представлен на всех глубинах бентального биотопа. В верхней части склона обычны зеленоглазки (род Chlorophthalmus), обитающие в широко батиметрическом диапазоне (от 100—150 до 1000 м). Эти небольшие (длина до 20 см) рыбы, которые ведут станую жизнь вблизи грунта и питаются бентосом. Как показано К. Судаком, к этому же семейству (Chlorophtha midae) принадлежат несколько родов, очень характерных для больших глубин (рис. 37). Батиптеры (около 20 видов рода Bathypterois длиной 15—30 см) встречаются в всех океанах на глубине 250—6000 м. Подводными наблюдениями и фотосъемкой установлено, что они обычно неподвижно стоят на дне, возвышаясь над ним, как на ходулях, на утолщенных и удлиненных краевых лучах брюшных плавников и хвоста (их называют еще рыбами-треногами), и выжидают появления планктонных рачков — их основной пищи. Длинные лучи грудных плавников выполняют при этом роль чувствительных антенн, так как глаза у этих рыб развиты плохо. В отличие от разнополых зеленоглазок батиптеры являются синхронными гермафродитами, как и другие глубоководные виды семейства — батимикропсы (Bathymicrops), батитифлопсы (Bathytyphlops) и ипнопсы (Ipnops), обитающие в батин абиссопелагиали. У представителей последнего рода глаза полностью редуцированы, а роль светочувствительных органов выполняют так называемые глазные пластины, расположенные на верхней поверхности головы под тонкими костями. Плодовитость батитифлопсов достигает, по Н. Мерритту, 6000 икринок, проходящих развитие в толще воды: их личинки, описанные как особый род Macristiella, живут в эпипелагиали. Иныопсы и батимикропсы откладывают не более 300 икринок, которые имеют отрицательную плавучесть. Эти рыбы, по-видимому, проводят у дна всю свою жизнь.
В близком семействе ящероголововых (Synodonlidae) большая часть видов живет на шельфе, хотя некоторые зауриды (род Saurida) встречаются и в верхней мезобентали — до глубины 550 м. Лишь два вида рода Bathysaurus относятся к числу настоящих глубоководных рыб — один из них (В. mollis) ловится преимущественно па глубине 2500—4500 м при температуре 2—3°. Батизавры — крупные хищные рыбы (длина до 85 см), питающиеся в основном рыбой. Они тоже синхронные гермафродиты, плодовитость составляет 24—56 тыс. икринок. На личиночно-мальковой стадии развития (ранее также относившейся к особому роду Macristium) держатся, по-видимому, в мезопелагиали. Особое место среди ящероголовов занимают придонно-пелагические бомбили — род Harpodon, из трех видов которого наиболее известна «бомбейская утка» (Н. nehereus) длиной до 30 см, широко распространенная в прибрежных водах Индийского и западной части Тихого океанов. В определенные сезоны эта рыба стаями появляется вблизи берегов и даже входит в опресненные эстуарий, где ее вылавливают в немалом количестве, но значительную часть жизни она, как предполагают, проводит на глубинах склона.
По внешнему облику бомбейская утка вполне соответствует представлению о глубоководной рыбе, и тем более неожиданной для меня была первая (и единственная) встреча с ней... В 1961 г «Витязь» уходил из Калькутты, крупнейшего порта Индии, расположенного на одном из рукавов дельты Ганга — реке Хугли, воду которой можно сравнить только с глинистым раствором: она имя ет цвет кофе с молоком и, подобно нашим среднеазиатским рекам несет огромное количество взвеси. На выходе в такой же мутный эстуарий к борту подошла небольшая парусная лодка с двумя рыбаками-бенгальцами, предложившими свой скромный улов несколько рыб, аккуратно уложенных в плетеную корзину. В этом то улове (мы получили его в обмен на сгущенное молоко) оказались бомбили. Бесчешуйные большеротые «утки» отличались дряблой консистенцией жирного тела, очень характерной для многих глубоководных рыб, и буро-коричневой окраской. Эти. «птицы», надо сказать прямо, не очень-то находят место в той экологической классификации, которая принята в этой книге - ведь из числа океанических рыб в эстуариях можно встретить, кроме них, пожалуй, только брегмацеров (один из них, кстати, был даже описан по экземпляру из эстуария Хугли).
Завершая обзор аулопообразных, надо отметить, что к числу бенто-пелагических рыб нужно относить и преобладающее большинство скопелозавров (род Scopelosaurus из сем. Notosudidae), взрослые особи которых держатся только над континентальными склонами, островными цоколями и подводными поднятиями.
Очень много донно-придонных океанических видов содержит отряд трескообразных рыб, две трети состава которого обитает на значительных глубинах. Преимуществ венно глубоководным является, в частности, семейство моровых (Moridae), впервые обособленное известным отечественным ихтиологом А. Н. Световидовым (1903—1985).
Судьба довольно близко свела меня с Анатолием Николаевичем Световидовым. Я познакомился с ним еще в середине 50-х годов, при одной из первых командировок в ленинградский Зоологическпй институт. Эту встречу помню хорошо. А. Н. пригласил меня в кабинет и, вскользь выяснив, кто я, откуда и чем интересуюсь, целиком взял нить разговора в свои руки и долго с большим увлечением говорил о своих студенческих годах (он учился на Отделении рыбоведения Московской сельскохозяйственной академии, из которого впоследствии образовался Мосрыбвтуз, так что, получалось, мы окончили один вуз), о своих старших коллегах учителях — В. К. Солдатове, Н. М. Книповиче и Л. С. Берге, которых глубоко чтил, о деталях своей тогдашней работы по черноморским рыбам. При последующих посещениях Ленинграда эти беседы-монологи вошли в обычай (видимо, я оказался хорошим слушателем), а через несколько лет я стал вхож и в квартиру на Московском проспекте (у А. Н. была прекрасная ихтиологическая библиотека) и на дачу в Комарове, познакомился с добрейшей Елизаветой Васильевной, стал иной раз останавливаться у Световидовых при коротких приездах (А. Н. всегда выговаривал мне, если я находил иное пристанище). С середины 70-х годов наши встречи стали очень регулярными: будучи членом бюро Отделения общей биологии Академии наук, А. Н. почти еженедельно бывал в Москве и постепенно у него вошло в привычку проводить у меня дома последние четыре-пять часов в ожидании полуночного курьерского поезда. Все это время он рассказывал, рассказывал и рассказывал...
Меня всегда поражали трудолюбие и организованность А. Н., который как бы сам задавал себе ежедневный урок и всегда выполнял его независимо ни от чего. Три капитальные монографии — «Сельдевые», «Трескообразные» и «Рыбы Черного моря», вошедшие в золотой фонд ихтиологии,— итог упорного многолетнего труда с полной отдачей сил и времени. Огромная требовательность к себе позволяла А. Н. быть чрезвычайно требовательным и к своим сотрудникам и ученикам, и он не прощал никому из них ни отступлений от генеральной линии исследований, ни отвлечений на посторонние, с его точки зрения, дела, ни тем более проявлений неусидчивости или недисциплинированности. Его отношения с коллегами по институту не были легкими, а с некоторыми из них он, оставив административную деятельность, не общался вовсе. По-видимому, именно поэтому он искал и находил себе собеседников на стороне и открывался им с самой неожиданной стороны — ведь его монологи касались не только профессиональных вопросов, но также истории, литературы, искусства. Впрочем, с любой темы А. Н. с неизбежностью возвращался к ихтиологии, которая, несомнено, была единственной настоящей страстью его жизни.
Моровые рыбы (их около 90 видов) близкородственны тресковым, но обитают в основном на глубинах материкового склона (в семействе есть, впрочем, и шельфовые виды, распространенные преимущественно в умеренных широтах южного полушария, а представители 3—4 монотипических родов живут, как предполагают, в толще воды). Они представлены во всех океанах и особенно многочисленны в верхней батиали, но некоторые виды встречаются в более широком диапазоне глубин — антимора (Antimora rostrata) (рис. 38), например, от 400 до 3000 м. Длина океанических морид варьирует от 15—25 см у физикулюсов (род Physiculus) до 80-90 см у лепидионои (Lepidion). Они сильно различаются по образу жизни.Физикулюсы, подобно налимам, ищут свою добычу на дне, пользуясь обонянием (поэтому они часто попадают в донные ловушки); антимора поедает как бентосных животных, которых она может выкапывать из мягкого грунта лопатообразным рылом, так и придонно-пелагических рыб и кальмаров; дальневосточная лемонема (Laemonema longipes) совершает суточные вертикальные миграции, поднимаясь ночью в пелагиаль, и питается там; макропланктоном, крупные лепидионы и моры (род Morа) едят креветок, рыб и кальмаров.
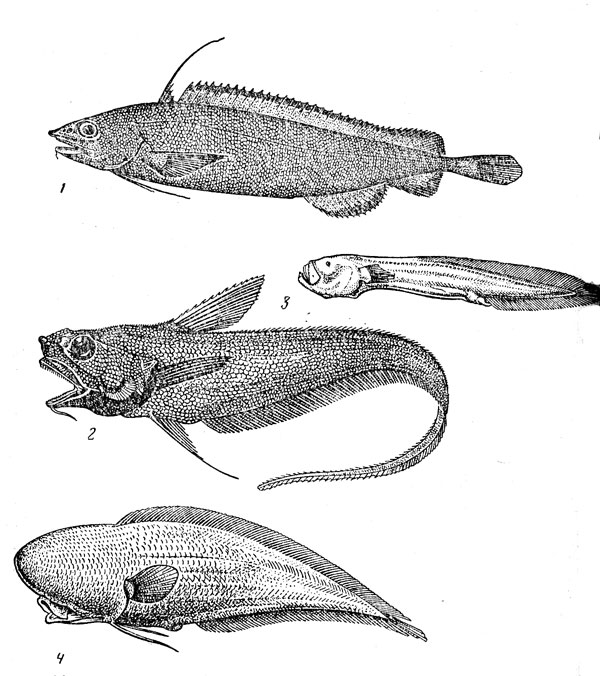
Рис. 38. Глубоководные трескообразные и ошибнеобразные рыбы: антимора (1) долгохвост (2), нибеллинелла (3), слепой тифлонус (4)
Биология глубоководных морид в целом изучена недостаточно. Исключение составляет, пожалуй, только лемонема, обитающая в северной части Тихого океана от берегов Хонсю до залива Аляска на глубине 400—1200 м. Лемонема — тугорослая рыба, созревающая при длине более 40 см в возрасте 5—8 лет и достигающая предельной длины около 70 см в возрасте 26 лет. Начальные годы жизни она проводит в толще воды, а затем опускается в придонные горизонты (по ночам мигрируя в пелагиаль), причем более крупные особи держатся в нижней части батиметрического диапазона, занимаемого видом. Дальневосточной лемонеме свойственны значительные сезонные перемещения: летом она концентрируется в основном на склонах восточного Хоккайдо и Курильских островов, а зимой уходит в воды Южной Японии, где нерестится, выметывая 300—800 тыс. мелких икринок, на глубине 600—800 м. Эта рыба служит объектом промысла.
Для самой верхней части склона, пограничной с шельфом, характерны также мерлузы, или хеки (род Merluccius), и макруронусы (Macruronus) — хищные рыбы, достигающие 0,7—1,3 м (все они имеют промысловое значение), из которых лишь немногие могут быть встречены вплоть до глубины 1000 м. Другие представители семейства Merlucciidae, например штайндахнерия (Steindaclmeria argentea) из западной тропической Атлантики, имеющая сложную систему светящихся органов, являются па-стоящими мезобентопелагическими рыбами.
Следующее семейство отряда трескообразных — длиннохвостовые, или макруриды (Macrouridae), — самая многообразная группа глубоководных рыб, насчитывающая более 300 видов. В последние годы она интенсивно изучалась В. М. Макушном, Н. Маршаллом, Т. Ивамото, Ю. И. Сазоновым, Д. Стейном, О. Окамура и другими исследователями. Макруриды широко распространены в Мировом океане (они отсутствуют только в Арктике) на глубине от 50—100 до 5—6 тыс. м. Чисто шельфовых видов в семействе, впрочем, очень немного, и все они принадлежат к роду полорылов — Coelorinchus). Естественно, отдельные виды имеют более узкое географическое и батиметрическое распространение. Преобладающее большинство долгохвостов приурочено к мезобентали и мезобентопелагиали (200—1500 м), причем многие из них в высшей степени стенобатны: у ряда видов незумий (род Nezumia) и гименоцефалов (Hymenocephalus) весь диапазон вертикального распределения укладывается в две-три сотни метров. Для нижней батиали (т. е. батибентали) особенно характерны роды Macrourus и Coryphaenoides (см рис. 38), а в абиссальную зону проникают лишь отдельные представители последнего (например, С. armatus), а также роды Echinomacrurus и Astheriomacrurus. Для верхнебатиальных видов очень типичен значительный региональный эндемизм (иначе говоря, они имеют ограниченное географическое распространение), но в более глубоких горизонтах ареалы долгохвостов, как правило, занимают обширные площади. Большинство видов обитает в тропической зоне.
Внешний вид долгохвостов чрезвычайно своеобразен: у них непропорционально большая голова и очень длинный постепенно утончающийся в нить хвост, за который англичане называют этих рыб «крысохвостами». Подобно отдельным моровым и мерлузовым рыбам, некоторые роды макрурид обладают одиночными или, реже, парными светящимися железами, расположенными на брюхе перед анальным отверстием и содержащими бактерий-симбионтов, излучающих свет в результате своей жизнедеятельности. Такая железа состоит из мешковидного резервуара для бактерий, светоотражающего слоя и прозрачной линзы. Она снабжается кровью через систему капиллярных сосудов, обеспечивающих бактерии кислородом, и светоиспускание может регулироваться как изменением интенсивности кровеснабжения, так и «зашториванием» линзы темными хроматофорами. Некоторые виды (например, мягкоголовый долгохвост Malacocephalus laevis), по-видимому, способны выпускать в воду струю светящейся жидкости.
Долгохвосты сильно различаются по размерам. Самые мелкие из них не превышают 15—20 см, а самые крупные достигают 2 м в длину. Среди них есть как настоящие донные формы, в поисках пищи перекапывающие грунт жесткими носами (жесткорылы рода Trachyrinchus и многие полорылы), так и питающиеся макропланктоном бентопелагические формы, составляющие явное большинство, а также немногочисленные чисто пелагические виды, которые упоминались в предыдущей главе. Все эти категории связаны переходами, так как многие долгохвосты могут быть встречены как на грунте, так и в отрыве от него. Некоторые виды из рода Coryphaenoides (тупорылый макрурус С. rupestris и другие) даже совершают регулярные миграции с подъемом в пелагиаль в ночноя время, но никогда не поднимаются в верхние слои воды
Более или менее полные представления об экологии существуют сейчас только в отношении тех долгохвостов, которые уже вовлечены в промысловое использование. К их числу относятся, в частности, северотихоокеапские малоглазый (Coryphaenoides pectoralis) и пепельный (С. cinereus) макрурусы, а также североатлантический тупорылый макрурус. Все они обитают в широком диапазоне глубин (от 140—180 до 2,5—3,5 тыс. м), но в массовом количестве встречаются только в верхней части материкового склона. Наибольшая длина этих видов 0,5—2,0 м, предельный возраст 10—40 лет. В соответствии с размерами находится и плодовитость: малоглазый макрурус откладывает до 400 тыс. икринок, тупорылый — до 60 тыс., пепельный — до 18 тыс. Несмотря на довольно высокую численность этих видов, их планктонные личинки попадаются в пробах очень редко, и местообитание ранних стадий развития остается неизвестным. Основную пищу всех трех видов составляет мезопелагический макропланктон — креветки, криль, кальмары, а из рыб — светящиеся анчоусы, значительно меньшая роль принадлежит бентосу, хотя малоглазый макрурус, например, почти не выходит в пелагиаль.
Хорошо представлено в глубинных бентопелагических комплексах семейство ошибневых (Ophidiidae), по видовому разнообразию уступающее одним только долгохво-стам. Сюда относится около 200 видов (достойный вклад в их изучение внесли И. Нильсен, Д. Коэн, Ю. Н. Щер-бачев), и почти три четверти из них ведут глубоководный образ жизни. Среди ошибневых есть верхнебатиальные формы, в частности мономитопы (род Monomitopus) и многие дикролены (Dicrolene), но особенно характерны они для нижней части склона и для ложа океана, где живет большинство видов семейства, в том числе наиболее мелкие (Tauredophidium hextii длиной немногим более 10 см) и наиболее крупные (Spectrunculus grandis длиной 1,3 м и массой 16,5 кг) его представители. К числу самых глубоководных видов принадлежат слепой тифлопус (Typhlonus nasus) (см. рис. 38), обнаруженный вплоть До глубины 5100 м, лентовидный бассоцет (Bassozetus taenia) — до 5600 м, атлантический лейцикорус (Leucicorus atlanticus) — до 6800 м, холкомиктеронус (Holcomycteronus profundissimus) — до 7160 м и абиссобротула (Abyssobrotula galatheae), которой принадлежит абсолютный рекорд по глубине вылова: в желобе Пуэрто-Рико она была добыта с глубины 8370 м. Все глубоководные офидииды во взрослом состоянии ловятся только донными тралами, но их личинки, по-видимому, живут в толще воды. Пищу этих рыб, нигде не достигающих особенно большой численности, составляют донные животные, в основном полихеты и ракообразные.
Сходный с офидиидами образ жизни ведут представители близкого к ним семейства бититовых (Bythitidae), которые отличаются способностью к живорождению (оплодотворение у них, естественно, внутреннее, и самцы имеют довольно сложно устроенный копулятивпый орган). Виды катетиксов (род Cataetyx), в частности, довольно обычны на всех глубинах континентального склона и встречаются во всех океанах. Взрослые особи этих видов живут на самом дне, а молодь, заметно отличающаяся от взрослых по внешнему виду, держится в толще воды. Бентопелагические афионовые рыбы (сем. Aphyonidae) длиной от 4 до 18 см — обитатели придонных горизонтов, встречающиеся в нижней батиали и над ложем океана (наибольшая глубина поимки — 5320 м ).Некоторые нетопыревые рыбы встречаются и гораздо глубже.
Следующий отряд — удильщикообразные — характерен преимущественно для мезобентали, в которой он представлен тремя семействами и примерно сотней видов. Большинство удильщиков (сем. Lophiidae), которых называют также морскими чертями, обитает на глубине до 500—600 м, но некоторые, например европейский морской черт (Lophius piscatorius), могут опускаться гораздо глубже - даже до 2000 м. Эти своеобразные рыбы длиной 0,3—1,2 мм имеют уплощенное тело, непропорционально большую голову и огромную зубастую пасть и проводят большую часть жизни, лежа на дне в пассивном ожидании добычи (в основном различных рыб), которую подманивают с помощью особой вымпеловидной приманки — эски, вывешенной перед ртом на конце «удилища» — длинного обособленного луча спинного плавника. В том же диапазоне глубин (200—500 м) обычны другие «удящие» рыбы, также относящиеся к числу бентических — жабовидные хаунаксы (сем. Chaunacinae) длиной до 40 см и морские нетопыри (Ogcocephalidae) (Так, голотип Halieutopsis andriashcvi — вида, недавно описанного из тропической зоны Индийского океана, был выловлен в батибентали на глубине около 4000 м.), редко превышающие 15—20 см в длину. Последние во взрослом состоянии практически теряют способность к плаванию: они перемещаются по дну только по-пластунски — ползком, используя для этого свои «руки» — грудные плавники, как бы согнутые в локтях, и «ноги» — брюшные плавники и лишь подталкиваясь небольшим хвостовым плавником. Все эти рыбы, по-видимому, откладывают икру на дно, но их личиночное развитие идет в толще воды. Вероятно, они имеют большую продолжительность жизни (один из некрупных видов морских нетопырей, к примеру, живет не менее 9 лет).
Среди преимущественно пелагических видов отряда опахообразных особое место занимают донные ателеоповые рыбы (сем. Ateleopodidae), внешне очень сходные с долгохвостами (их истинное место в системе было установлено в самое недавнее время). Некоторые из них (представители рода Ijimaia) достигают крупных размеров (длина до 2 м) и широко распространены в тропической зоне на глубине от 200 до 1000—1500 м, хотя повсюду относятся к числу редких рыб.
«Книжное» — по описаниям и рисункам — знание рыб, конечно, позволяет узнавать этих рыб при встрече, но их истинный облик может очень сильно отличаться от того представления, которое сложилось до личного знакомства. Должен сказать еще, что мы, ихтиологи, обычно различаем уже известные нам виды не по их официальным диагностическим признакам, а по общему облику (как говорится, габитуально), т. е. так, как это делают все рыбаки (многие из них, пожалуй, не смогут ответить на вопрос, чем отличается лещ от густеры, но опознают их вполне уверенно).
С ателеопом мне довелось впервые встретиться совсем недавно - только в мае 1987 г., когда «Профессор Штокман» проводил исследования на подводном хребте Сала-и-Гомес. Естественно, я вполне определенно представлял себе эту уникальную рыбу, но действительность превзошла все ожидания. Увидев в развязанном куте трала среди ярких колючих антигоний, тригл и окуней белесый свинцового цвета глаз с крохотным темным окошечком в центре, я сразу обратил на него внимание. Глаз был окружен чем-то гладким и студнеобразным, и я только успел подумать об осьминоге, как Юрий Сазонов поставил диагноз: «Ателеопус попался!» (он уже видел этих рыб, работая на траулере в Индийском океане). Три наших атолеопа, пойманные па глубине 380 м, имели чуть больше полуметра в длину. Покрытые гладкой скользкой кожей, они больше всего- напоминали огромных головастиков. Наибольшее впечатление производила их голова — совершенно не рыбья, мягкая и полупрозрачная как желе, обтянутая нежной скользкой кожей. Маленький, воронкообразный и совсем беззубый ротик, расположенный на нижней поверхности, вызывал сильное сомнение в способности его обладателя питаться «рыбой, офиурами и ракообразными» (именно эти объекты находили в желудках ателеопид). По необычности облика ателеопы нисколько не уступают ни удильщикам, ни хаулиодам, ни мешкоротам.

Рис. 39. Глубоководные бериксообразвые рыбы: стефаноберикс (1), средиземноморский хоплостет (2), диретмоид (3), низкотелый берикс (4).
Существенная роль в ихтиоценах материкового склона и подводных поднятий принадлежит некоторым представителям отряда бериксообразных рыб (рис. 39). Наиболыним разнообразием среди них выделяется семейство трахихтовых (Trachichthyidae), детально изученное в последние годы А. Н. Котляром. Почти все представители этой группы (а их около 40) относятся к числу полуглубоководных рыб, весьма обычных в нижней части шельфа и над его свалом на глубине 150—800 м. Один из наиболее широко распространенных видов — атлантический большеголов, или пилобрюх (Hoplostethus atlanticus), встречается у обоих берегов Атлантического океана и в умеренно теплых водах южного полушария на глубине от 300 до 1500 м (чаще всего 800—1200 м). Этот вид, как и другие трахихтовые, характеризуется медленным ростом (годовые приросты порядка 2—3 см) и большой продолжительностью жизни — до 24 лет. Половой зрелости он достигает в возрасте 10—15 лет (плодовитость варьирует от 20 до 200 тыс. икринок), а наиболее крупные особи имеют длину около 60 см при массе около 4,5. Его пищу составляют макропланктонные животные: ракообразные, мелкие кальмары и рыбы.
В придонных слоях воды над материковым склоном и подводными поднятиями (обычная глубина 200—800 м) обитают бериксовые рыбы (сем. Berycidae). Низкотелый берикс (Beryx splendens) — ярко-красная стайная рыба, внешне напоминающая морского окуня и достигающая 45 см в длину, встречается во всех океанах. В дневное время он держится над дном, а ночью отрывается от него и выходит в водную толщу, нередко образуя значительные скопления. Пищу берикса составляет различный макропланктон, преимущественно ракообразные, рыбы (в том числе миктофиды), мелкие кальмары.
Из других придонных бериксообразных нужно отметить диретмоидов (род Diretmoides), которые вместе с мезопелагическим диретмом (см. гл. III) входят в состав семейства Diretmidae. Они достигают 40—50 см в длину, окрашены в темные тона и во взрослом состоянии постоянно пребывают над дном на глубине порядка 500— 800 м, хотя их серебристые мальки, очень похожие на диретма, ведут пелагический образ жизни. В нижней батиали живут также гиспидоберикс (Hispidoberyx ambagiosus), совсем недавно (в 1980 г.) описанный А. Н. Котляром и выделенный им в особое семейство, и гибберихты — два вида рода Gibberichthys. Личинки последних при длине 1—2 см резко отличаются по внешнему виду от взрослых рыб и до последнего времени считались принадлежащими к самостоятельному семейству. У этих рыбок сильно удлиненный третий луч брюшных плавников имеет вид длинного стержня, несущего около 30 больших листовидных мешков черного цвета с прозрачным овальным окошечком па конце каждого. Эти придатки внешне очень напоминают некоторых сифонофор или медуз с их плавательными пузырями и свисающими щупальцами и могут расцениваться как проявление мимикрии (не исключено, впрочем, что они испускают свет и служат для приманивания добычи). Личинки и мальки гибберихтов держатся в поверхностных слоях воды, взрослые рыбы живут в придонных горизонтах на глубине 750—2000 м.
Среднеглубинные бентопелагические рыбы есть и в отряде солнечникообразных. Одни из них — антигонии (род Antigonia) и солнечники (Zeus и Zenopsis) имеют центром батиметрического распределения зону, переходную между эпи- и мезопелагиалью (глубины порядка 100—400 м), другие, принадлежащие к семействам ореосомовых (Oreosomatidae) и чешуеиглых (Grammicolepididae), встречаются только в верхней батиали — на глубине от 200—500 до 1500—2000 м. Аллоцит, или лунник (Allocyttus verrucosus), длиной до 38 см широко распространен в Атлантике и южной части Индийского океана и кое-где (например, на банке Агульяс) даже образует плотные концентрации. Питается эта рыба разнообразным макропланктоном.
Бенто-пелагический образ жизни в диапазоне глубин от 100—200 до 600 м ведут морские бекасы (сем. Масrоrhamphosidae), принадлежащие к отряду колюшкообразных. Наиболее обычный вид — морской бекас (Масrоrhamphosus scolopax) длиной до 19 см — широко распространен в тропических и субтропических водах и передко встречается в огромном количестве и очень плотными стаями над относительно мелководными подводными поднятиями. Считается, что молодые бекасы, живущие в отрыве от дна, имеют более прогонистое тело и окрашены с преобладанием синих тонов, тогда как у взрослых тело выше, а в окраске больше красного цвета, но эта точка зрения не общепринята (Т. Кларк считает, что существуют два вида этих рыб).
Глубоководные виды имеются в ряде семейств скорпенообразных рыб, более характерных для прибрежной зоны. В своем преобладающем большинстве они приурочены к самым верхним горизонтам мезобентали, пограничным с эпибенталью. Вплоть до глубины 300—500 м (иногда и более) вполне обычны, в частности, некоторые скорпеновые рыбы (сем. Scorpaenidae) — морские окуни (Sebastes), беспузырники (Helicoienus) и др. Все они во взрослом состоянии ведут бентический образ жизни (молодь встречается, однако, в толще воды) и редко отрываются от дна, но из этого правила есть исключения. Суточные вертикальные миграции с ночным подъемом к поверхности отмечены у многих видов, а североатлантический окунь-клювач (S. mentella) вполне способен существовать в пелагиали, образуя значительные скопления даже над океаническими глубинами. Из тропических видов наиболее «пелагизирован» батиально-пелагический Ectreposebastes imus, имеющий в отличие от других среднеглубинных скорпенид черную, а не красную окраску. К настоящим мезобеитальным рыбам принадлежат, однако, немногие скорпеновые, постоянно живущие на глубине порядка 1000—1600 м («рекордная» глубина 2500 м) — шипощеки (род Sebastolobus), обитающие в северной части Тихого океана, и трахискорпии (Trachyscorpia). Размеры глубоководных скорпенид от 15—20 см до 1 м (обычно не крупнее 50 см). Все они, по-видимому, относятся к числу медленнорастущих, долгоживущих (до 10— 25 лет) и поздно созревающих рыб с невысоким темпом воспроизводства.
Для вод материкового склона характерно также семейство аноплопомовых (Anoplopomatidae). Угольная рыба (Aiioplopoma fimbria), достигающая длины 1,2 и массы 14 кг, обитает в северо-восточной Пацифике (от Калифорнии до Аляски), где встречается на всех этапах жизненного цикла. Есть она и в Беринговом море (там, по-видимому, существует зависимая популяция вида), а отдельные особи изредка ловятся даже у Курильских островов и Японии. Угольная рыба обитает на глубине 200—1200 м (максимальная глубина вылова 1800 м) и адаптирована к характерной для этих глубин пониженной концентрации растворенного кислорода. Личинки и мальки держатся в эпипелагиали, годовики длиной 20—40 см переходят к жизни у дна. Главную пищу составляют рыба, головоногие моллюски и кишечнополостные (гребневики и медузы). Второй вид семейства — эрилепис (Erilepis zonifer), также обитающий в северной части Тихого океана, принадлежит, вероятно, к верхне-мезобенто-пелагическому ихтиоцену.
Несколько мезобентальных видов содержит холодноводное семейство рогатковых (Cottidae), н все они имеют некрупные размеры (не более 15—20 см). Так, обычные на севере Тихого и Атлантического океанов и в морях Арктики ицелы (род Icelus) и триглопсы (Triglops) встречаются у дна вплоть до глубины 500—900 м, а самый глубоководный представитель группы — дальневосточный крючкорог Zesticelus profundorum — ловился даже на глубине свыше 1000 м. На материковом склоне живут и австрало-новозеландские рогатки-антиподы (род Antipodocottus), а также единственный экваториальный вид группы — Stlengis mesembrinus, описанный недавно из вод Индонезии. В близком семействе психролютовых (Psychrolutidae) все виды живут за пределами шельфа (общий батиметрический диапазон 150—1600 м). Некоторые роды (например, Psychrolutes и Cottunculus) имеют биполярное распространение.
Семейство липаровых, или морских слизней (Liparididae), очень богатое видами (их более 150), включает как прибрежные формы, обитающие только за пределами теплых вод, так и глубоководные виды, представленные во всех широтных зонах Мирового океана. Название «морские слизни» очень точно характеризует внешний облик этих рыб, голое тело которых имеет студнеобразную консистенцию. Их обычная длина составляет 10—30 см (максимально 70 см). В мезо- и батибентали особенно обыкновенны роды карепроктов (Careproctus) и паралипарисов (Paraliparis), каждый из которых насчитывает примерно по 60 видов. Карепрокты обитают на глубине до 3500—4000 м, причем шельфовые виды имеют светлую или ярко-розовую окраску, батиальные — красную или фиолетовую, а абиссальные — почти черную. В соответствии с глубиной обитания меняется также цвет брюшины (от светлой до черной) и диаметр глаз (наибольшего размера они достигают у видов, живущих в сумеречной зоне — на глубине 200—1000 м). Паралипарисы, обитающие в диапазоне глубин от 125 до 3200 м, наиболее многообразны в батиали северной Пацифики и, как показал А. П. Андрияшев, в Южном океане, где находится вторичный центр видообразования группы. Среди липарид имеются даже хадобентальные виды: родственный кapeпроктам псевдолипарис (Pseudoliparis amblystomopsis) обнаружен на глубине около 7600 м в Курило-Камчатском желобе, а три вида рода Notoliparis в трех разных желобах южного полушария (Кермадекском, Южно-Сандвичевом, Макуори) на глубине более 5000 м.
Мне не приходилось всерьез заниматься липаридами, по все же у меня есть одно воспоминание, правда, отнюдь не из приятных, связанное с этими рыбами. В 1979 г. «Витязь» обогнул Европу в своем последнем 65-м рейсе, посетив о «прощальным визитом» порты Франции, Испании, Португалии, Англии и Дании. В том рейсе принимал участие и А. П. Андрияшев (мы жили в одной каюте), который очень надеялся использовать посещение Англии для изучения типовых экземпляров антарктических паралипари-сов в Британском музее естественной истории. В Бискайском заливе нас накрыл жестокий шторм, и Анатолий Петрович, поскользнувшись на трапе в момент, когда судно резко повалило на борт, получил очень тяжелую травму — перелом нескольких ребер, в результате чего оказался в больнице города Дувра. О визите в Лондон нечего было и думать, и он попросил меня проверить некоторые признаки интересовавших его рыб попутно с моим исследованием типового материала по летучкам. Для работы в музее (я ездил туда два раза) мне выделили отдельную комнату, куда были доставлены из хранилища банки с отобранными по каталогу экземплярами. Покончив с летучими рыбами, я открыл сосуд с паралипарисами. Три сиптипа P. antarcticus - вида, описанного Т. Ригеном еще в 1912 г., находились в неважном состоянии: вероятно, они были исходно фиксированы спиртом, сильно дегидратировались и стали очень твердыми. Пытаясь распрямить одну из рыбок для измерения, я сделал слишком резкое движение и, о ужас, обломил ее хвост. Я оцепенел: изуродовать типовой экземпляр — это непростительное преступление, которому но может быть прощения. Но делать нечего... С повинной головой я направился к Питеру Уайтхеду - куратору коллекции рыб и покаялся в своем грехе. «А где хвост?» - спросил Питер. «Вот он». - «Ну, ничего. Мы его подклеим так, что будет совсем незаметно». Я был прощен, по до сих пор испытываю чувство стыда, вспоминая это событие.
Самый большой отряд рыб — окунеобразные — содержит свыше 6000 видов, но лишь очень немногие из них относятся к глубоководным обитателям дна и придонных слоев пелагиали, а на абиссальные глубины вообще проникают считанные формы. Сколько-нибудь значительную роль они играют, пожалуй, только в пограничной зоне между шельфом и склоном, где встречаются многие преимущественно шельфовые виды каменных окуней (сем. Serranidae), каталуфовых (Priacanthidae), бланквиловых (Blanchiostegidae), мицуевых (Scombropidae), красноглаз-ковых (Emmelichthyidae), рыб-кабанов (Pentacerotidae) и Других семейств, не опускающиеся, как правило, глубже 400-600 м.
Только для мезобентопелагиали характерны семейства акропомовых (Acropomatidae) и эпигоновых (Epigonidae), к которым принадлежат рыбы, в преобладающем большинстве живущие в придонных горизонтах на глубине 200—600 м. В обоих семействах есть, однако, виды которые населяют более глубокие слои (800—1100 м), например хауэлла (Howella brodiei), широко распространен пая во всех океанах, и южный эпигонус (Epigonus robustus), населяющий материковые склоны и подводные поднятия в 30—40-х широтах южного полушария. Один из наиболее крупных видов — эпигонус-телескоп (Е. telescopus), достигающий длины свыше 70 см и массы 4 км также обитает на глубине до 1200 м, где является объектом промысла. Мальки многих эпигонусов держатся в верхнем слое океана.
Интересно отметить, что в преимущественно эпимезопелагическом семействе морских лещей брамид виды одного рода — Eumegistus, являющегося, по мнению Д. Мида, наименее продвинутым в эволюционном отношении, ведут придонный образ жизни и выходят в толщу воды только для ночной охоты на миктофид и подобную им по поведению добычу. Эти лещи достигают довольно крупных размеров (до 70 см) и живут над островными склонами и подводными горами в тропической зоне.
Бельдюговые (сем. Zoarcidae) — очень многообразная холодноводная группа северного происхождения, насчитывающая в своем составе около 200 видов, большая части которых обитает на глубине 100—500 м и даже глубже, хотя в семействе есть и мелководные, в том числе литоральные, формы. Почти все бельдюговые живут непосредственно на дне и тяготеют к мягким илистым грунтам в которые способны зарываться сами и закапывать свою икру; питаются они бентосом и донными рыбами. Исключение в этом отношении составляют только меланостигмы (род Melanostigma) и ликодапусы (Lycodapus) — они могут считаться мезобентопелагическими, так как нередко встречаются в значительном удалении от дна над большими глубинами.( Аналогичный образ жизни ведут, по-видимому, представителя близкого семейства Parabrotulidae — мелкие (до 5 см) живоротдящие рыбки двух родов (Leucobrotula и Parabrotula), ) Эти рыбы размножаются, однако как и все их родичи, откладывая крупную донную икру (прямых наблюдений нереста, впрочем, не проводилось). В отличие от преобладающего большинства других глубоководных рыб ранние мальки этих и всех остальных зоарцид в верхних слоях пелагиали не отмечены.
В изучение океанических зоарцид, принадлежащих главным образом к подсемейству ликодоподобных (Lycodinae), большой вклад внесен в последние годы А. П. Андрияшевым, В. В. Федоровым, Э. Андерсоном. К обширному (около 50 видов) роду ликодов (Lycodes) относятся в основном крупные (длина до 1 м) верхне-мезобентальные формы, и лишь немногие его представители доходят до батибентали. Близкие к ликодам, но более миниатюрные лиценхелисы (род Lycenchelys) заметно более глубоководны: эти рыбы полностью отсутствуют па шельфе и входят в состав мезо-, бати- и абиссобентальных ихтиоценов. Оба рода богато представлены во всех северных морях (они обычны и в наших дальневосточных и полярных водах), но встречаются и в южном полушарии, куда их предки проникли с севера по глубинам. В умеренно холодных водах Южной Америки они, поднявшись на шельф, дали начало эндемичным южным родам зоарцид, обитающим и на мелководьях. К числу наиболее глубоководных зоарцид принадлежит и пахикара (Pachycara obesa) (рис. 40), обитающая в Атлантическом океане на глубине 2400—4800 м. Близкий род Thermarces был описан совсем недавно Р. Розеиблаттом и Д. Коэном из термальных источников в восточной части Тихого океана на глубине 2600 м. В желудках термарцесов были найдены остатки моллюсков, раков амфипод и вестиментифер рнфтий, существующих за счет сульфобактерий, что позволяет считать их постоянным компонентом этих своеобразных глубоководных сообществ.
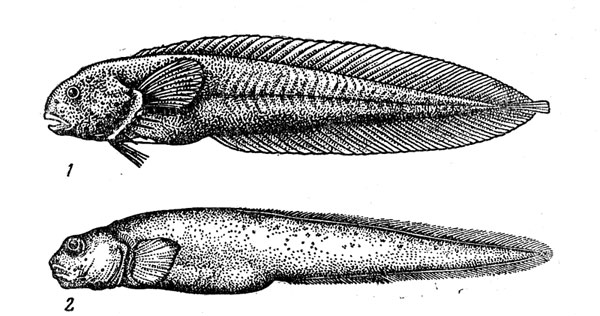
Рис. 40. Паралиприс (1) и пахикара (2).
Зубатки (два рода семейства Anarhichadidao), распространенные в северных частях Атлантического и Тихого океанов, живут в основном в шельфовой зоне, но в зимнее время мигрируют на глубины (эта группа всесторонне изучена В. В. Барсуковым). Наиболее глубоководный вид - синяя зубатка (Anarhichas denticulatus), достигающая 1,4 м в длину, отмечена даже на глубине более 900 м. Все зубатки откладывают очень крупную донную икру, но их личинки поднимаются в толщу воды, где проходят весь мальковый период жизни (его продолжительность у разных видов сильно разнится).
Еще одна холодноводная группа рыб — нототениевидные, эндемичная для Антарктики и Субантарктики, где она доминирует. Эта группа отличается, как показано работами А. П. Андрияшева, X. Девитта, А. В. Балушкина, А. В. Неелова, Ж.-К. Юро и других ученых, большим систематическим и экологическим разнообразием. В сем. нототениевых (Nototheniidae), усатых дракончиков (Artedidraconidae), антарктических плосконосов (Bathydraconidae) и белокровных рыб (Channichthyidae) есть и глубоководные донно-придонные формы, обитающие на глубине 500—1500 м и более. Среди преимущественно прибрежных и шельфовых нототениид наиболее глубоководными являются антарктическая сквама (Lepidonotothen kempi) и черноротый трематом (Trematomus loennbergi), которые держатся на склоне континентальной ступени Антарктиды на глубине 500—1000 м, а также патагонский клыкач (Dissоstichus eleginoides) — очень крупная рыба (длина до 2,1 м, масса до 70 кг), добываемый у островов Южная Георгия и Кергелен на глубине до 800 м. Большинство плосконосов живет на глубине, не превышающей 500—800 м, но представители рода Bathydraco — самого глубоководного среди нототеноидных рыб — населяют большие глубины (вплоть до 2500 м). Усатые дракончики рода Pogonophryne также отмечены до глубины 2300 м. Глубинная белокровка (Chionobathyscus dewitti), у которой, как и у других ханнихтид, в крови нет ни эритроцитов, ни гемоглобина, также ловилась у берегов ледового континента на глубине до 2000 м. У всех нототениевидных икра развивается на дне а личинки ведут пелагический образ жизни.
В верхней мезобентали тепловодной зоны (на глубине до 500—600 м) вполне обычны также такие рыбы, как хампсодоны (сем. Champsodontidae), звездочеты (Uranoe scopidae: Kathetostoma, Uranoscopus и др.), перкофид (Percophidae, в том числе карликовые виды родов Osopesaron и Pteropsaron, не превышающие 4—5 см в длину) драконетки (Draconettidae).

Рис. 41. Руветта (1) а угольная рыба-сабля (2).
Для среднеглубинных горизонтов очень характерны саблеподобные рыбы (рис. 41) — два близкородственных семейства гемпиловых (Gempylidae) и волосохвостых (Trichiuridae), входящие в состав подотряда скумбриевидных. Все они имеют в той или иной степени удлиненное тело, большой рот и очень острые мощные клыкообразиые зубы, легко рассекающие добычу на части. В мезобентопелагических ихтиоценах на глубине 300—1000 м (нужно сказать, что эта группа представлена также на шельфе и в океанской пелагиали) особенно важное значение принадлежит прометихтам (род Promethichthys), рексиям (Rexea), бентодесмам (Benthodesmus) и угольным саблям (Aphanopus) — хищным рыбам, длина которых нередко достигает 1 м, а также еще более крупной руветте (Ruvettus pretiosus). Эта рыба может иметь длину 1,8 м при массе около 50 кг. Во многих районах мира она служит объектом специального лова на особые снасти, крючки которых опускаются на глубину 300—700 м. Такой промысел производится, в частности, у берегов Кубы, Мадейры, Коморского архипелага, многих островов Полинезии. Мясо руветты содержит очень много жира, который обладает слабительными свойствами (на некоторых языках ее называют «касторковой рыбой») и при приготовлении рыбы в пищу должен удаляться во время варки.
Саблеподобным рыбам я посвятил немало внимания и времени. Еще в 1955 г., в первом своем плавании в тропиках, мне удалось поймать сачком крупную змеиную макрель, в то время представлявшуюся большой редкостью. В очень популярной в те годы книге Тура Хейердала о путешествии на бальсовом плоту «Кон Тики» встрече с этим «чудищем» посвящено немало эмоциональных строк. С тех давних пор эта группа постоянно вызывала мой интерес, и в 1969 г. я предложил Владимиру Беккеру, давнему приятелю (мы вместе учились в институте) и коллеге по лаборатории, совместно разобраться в не очень большой коллекции, которой мы тогда располагали: я взялся за рыб-сабель, он — за гемпилид. В итоге этого внепланового исследования был опубликован предварительный обзор надсемейства, после чего Володя навсегда вернулся к своим любимым миктофидам, а я тоже занялся другими проблемами, хотя не терял группу из виду и между делом описал в ней еще три вида бентодесм. В середине 70-х я заинтересовал систематикой трихиурид промыслового ихтиолога из ВНИРО Сергея Михайлина, вместе с которым мы дали описания двух новых рыб-сабель, но трагическая гибель Сережи в конце 1981 г. не позволила ему до конца реализовать свои возможности (он погиб, как герой, спасая пассажиров в загоревшемся на ходу вагоне поезда Калининград - Москва). В память о друге (нас объединяло не только общее дело, но и одинаковая страсть к охоте) я назвал его именем один из новых видов угольных сабель, решив поставить на этом точку в изучении этой группы. Не получилось... При обработке коллекций с подводного хребта Наска выяснилось, что там обитают два вида рексий - оба неописанные,— а исследование сравнительных материалов из других районов Тихого и Индийского океанов (я получил их из музеев многих стран) показало, что в роде существуют по меньшей мере шесть видов (считалось, что их только два). Результаты этой работы еще не опубликованы.
Из океанических строматеевидных рыб к числу придонных принадлежат немногие. Это — гипероглифы (род Hyperoglyphe) и шедофилы (Schedophilus) из семейства центролофовых — крупные рыбы длиной до 1 м, обитающие главным образом в субтропических и умереннотепловодных районах (в том числе над подводными поднятиями) на глубине до 300 м и более. Придонно-пелагическими; являются также ариоммы (около полутора десятков видов рода Ariomma), обычные на глубине 100—600 м во всех тропических морях.
Не опускаются глубже мезобентали и преимущественно шельфовые камбалообразные рыбы. Так, наиболее глубоководпый род ботусовых камбал (сем. Bothidae) — хасканопсетты (Chascanopsetta), недавно ревизованный К. АмаЯ ока, содержит 5 видов, которые живут на глубине 250-640 м. Один из этих видов — гавайская пеликанья камбала (Ch. crumenalis) отличается крайне своеобразным обликом: нижняя челюсть у нее очень длинная (она на треть: своей длины выступает перед рылом) и поддерживает кожистую горловую сумку, очень напоминающую ту, что имеется у настоящих пеликанов. Еще глубже обитают представители эндемичного для Южного океана семейства безруких камбал (Achiropsettidae), недавно выделенного С. А. Евсеенко и характеризующегося отсутствием грудных плавников: максимальная глубина вылова неоахиропсетты (Neoachiropsetta milfordl) составляет 840 м.
Среди настоящих, или правосторонних, камбал (сем. Pleuronectidae) есть немало видов, например северные малороты (род Microstomus) или тропические пецилопсетты (Poecilopsetta), приуроченных к пограничной между шельфом и склоном зоне (глубина порядка 100— 300 м). К наиболее глубоководным видам относятся палтусы, прежде всего черный, или синекорый (Reinhardtius hippoglossoides), и белокорый (Hippoglossus hippoglossus) — живущие в северных частях Атлантического и Тихого океанов хищники, активно преследующие свою добычу в толще воды. Полноразмерные особи черного палтуса достигают 1,3 м в длину, но белокорый значительно крупнее (рекордная длина 4,7 м при массе 337 кг). Скопления всех палтусов обычно приурочены к материковому склону (глубина 150—800 м), а наибольшая глубина их обитания превышает километр (черный палтус вылавливался даже па отметке 1700 м). Палтусы принадлежат к числу долгоживущих поздносозревающих рыб (белокорый живет до 30 лет, а созревает в массе на 10—15-м году). Нерест происходит в глубоких слоях воды, личинки сначала тоже держатся далеко от поверхности, но, подрастая, перемещаются на мелководья, где переходят к донному образу жизни. Среди камбал-плевронектид известны и еще более глубоководные — мезобатибентальные виды, например бородавчатая (Clidoderma asperrimurn) и глубинная (Еmbassichlhys bathibius), вполне обычные вплоть до 1800— 2000 м от поверхности. В состав батиального ихтиоцена полноправно входят и морские языки из рода Symphurus (сем. Cynoglossidae), живущие в тропической зоне на глубине 200—1000 м. К сказанному о камбалообразных нужно еще добавить, что среди глубоководных представителей группы практически нет чистых бентофагов (и уж во всяком случае их доля значительно ниже, чем среди мелководных).
И наконец, в отряде скалозубообразных к придонным глубоководным рыбам относится в полном составе семейство холлардиевых (Triacanthodidae), включающее 20 видов, обзор которых сделан недавно Д. Тайлером. Эти небольшие (длина не более 25 см) рыбы живут в тропических водах всех океанов на глубине 50—600 м и даже глубже. Биология их изучена слабо. Судя по некоторым особенностям, это наименее продвинутая группа скалозубообразных, но отдельные виды представляются очень сильно специализированными. Так, макрорамфосоды (род Macrorhamphosodes) имеют тонкое рыло, вытянутое в длинную трубку, на конце которой находится обращенный вбок дисковидный рот. Эта рыба, как и тидемания (Tydemania navigatoris), питается чешуей других рыб.
Сравнение состава бенто-пелагических комплексов разных вертикальных зон (табл. 4) показывает, что число видов в ихтиоценах быстро убывает с глубиной (цифры этой таблицы следует рассматривать как приблизительные). В самой верхней части склона, где представлены многие семейства, типичные для сублиторали, наблюдается наибольшее разнообразие систематического состава. Нужно подчеркнуть, что в большинстве случаев речь идет не о случайных проникновениях отдельных особей тех или иных шельфовых видов в чуждый биотоп, а о постоянном их обитании в экотонных условиях у границы смежных вертикальных зон. В справедливости такого утверждения убеждает тот факт, что такие виды живут и там, где эпибентальной зоны нет вообще. Речь идет о подводных поднятиях в открытом океане, имеющих минимальные глубины порядка 200—400 м, т. е. о верхней части талассобатиали в понимании А. П. Андрияшева. Все такие поднятия (их обычно называют «банками») населены тем не менее рыбами, которые более характерны для нижней части шельфа: как правило, их легко сразу опознать по яркой окраске и совсем не «глубоководному» облику. В качестве типичных эпимезобентальных экотонных рыб можно упомянуть аулопов (род Aulopus), длиннорылых рыб-солдатов (Ostichthys), антигоний (Antigonia), большеглазых каталуф (Cookeolus), каллантиасов (Callanthias). Верхнемезопелагические виды недавно обнаружены даже в семействах щетинозубовых, или рыб-бабочек (Chaetodontidae), и губановых (Labridae), всегда считавшихся приуроченными к прибрежным сообществам, — Chaetodon guyotensis и Bodianus thoracotaeniatus описаны с подводных гор хребта Кюсю-Палау (глубина 320—340 м).
На сообществах рыб, населяющих подводные поднятия, нужно остановиться подробнее в связи с тем интересом, который они вызывают по многим причинам. Важность их изучения первым оценил американский ихтиолог К. Хаббз, который еще в 1959 г., основываясь на очень скудных фактических данных того времени (фактически на одной случайной поимке триглы на гайоте хребта Наска), сформулировал ряд принципиальных вопросов, касающихся этой проблемы. Вот эти вопросы.
Постоянен ли видовой состав населения отдельных гор Каким образом они были заселены?
Не представляют ли они опорные пункты для преодоления открытоводных барьеров, разделяющих прибрежные фауны?
Не являются ли некоторые виды, населяющие постепенно погружающиеся горы, реликтами, адаптированными к увеличенной глубине обитания?
Ведет ли изоляция гор к видообразованию?
Не существует ли на горах и банках и над ними скоплений донных или пелагических рыб и других промысловых объектов?
Карл Левитт Хаббз (1894-1979) несомненно был одним из самых выдающихся ихтиологов нашего столетия. Он отличался необычайной широтой и глубиной научных интересов и оставил после себя весьма достойное наследие - более 700 книг и статей, касающихся разнообразнейших аспектов ихтиологии и смежных отраслей знания. Он был также простым в общении и доброжелательным человеком с хорошо развитым чувством юмора.
Мне довелось встретиться с К. Хаббзом трижды (не могу запросто называть его Карлом, как при встречах: по-русски это было бы слишком фамильярным и не соответствующим тому уважению, которое я питаю к этому человеку). Знакомство состоялось на Гавайских островах в 1964 г. во время очередного Тихоокеанского научного конгресса, на который я попал с «Витязем», совершавшим рейс в центральную Пацифику. Первое общение было очень тяжелым как по причине моего несовершенного английского, так и из-за весьма далекого от классических оксфордских норм произношения К. Хаббза, по я мог с удивлением убедиться, что даже в очень узкой специальной области (речь шла о летучих рыбах Тихого океана) он чувствовал себя совершенно свободно. На следующий день после моего доклада он шутливо поздравил меня с победой в противоборстве «с этим варварским английским», а через пару месяцев прислал в Москву вырезку из какого-то комикса, в котором говорилось о «летучих коровах», выдуманных неким впоследствии разоблаченным лжецом, написав на ней: «А существуют ли все же летучие рыбы?» В 1968 г. «Академик Курчатов» зашел в Виллемстад (о-в Кюрасао), когда там происходила международная конференция по Карибскому морю, в которой, оказалось, принимал участие и К. Хаббз. Он был очень подвижек и каждый год предпринимал дальние вояжи вместе со своей верной женой и помощницей Лорой Хаббз. На конференции он заметил меня первым (я, надо сказать, совсем не ожидал этой встречи), сразу узнал и с такой непринужденностью «проехался» насчет моей изменившейся внешности (я стал носить бороду), что два молодых ихтиолога с Барбадоса, с которыми я беседовал, с полной уверенностью сочли его моим бывшим профессором. Последний раз я видел Хаббза в 1979 г. в Калифорнии. Он был болен и знал, что его дни сочтены, сильно ослабел и постарел, но продолжал трудиться, интересовался работой коллег, говорил о желании принять участие в следующем Тихоокеанском конгрессе в Хабаровске... Прах Карла Хаббза по его завещанию был развеян над Тихим океаном. Большой фотопортрет с надписью «...советскому коллеге от старого моряка», запечатлевший его стоящим в полный рост на корме штормующего в океане судна, напоминает мне о наших встречах.
Таблица 4
| Отряд, семейство | Всего видов | Видов в ихтиоценах | Встречаемость в других ихтиоценах | |||||||
| ЭМ | М | МБ | Б | БА | А | АХ | Х | |||
| Миксинообразные (Myxiniformes) | ||||||||||
| Миксиновые (Myxinidae) | 32 | 20 | 6 | 1 | - | - | - | - | - | Эпибенталь |
| Многожаберникообразные (Hexanchiformes) | ||||||||||
| Плащеносные акулы (Chlamydoselachidae) | 1 | - | 1 | - | - | - | - | - | - | Нет |
| Катранообразные (Squalijormes) | ||||||||||
| Катрановые акулы (Squalidae) | 68 | 12 | 38 | 8 | - | 1 | - | - | - | Эпимезопелагиаль |
| Центриновые акулы (Oxynotidae) | 4 | 2 | 2 | - | - | - | - | - | - | Эпимезопелагиаль |
| Ламнообразные (Lamniformes) | ||||||||||
| Песчаные акулы (Odontaspidae) | 4 | 1 | 1 | - | - | - | - | - | - | Эпибенталь |
| Акулы-домовые (Mitsukuriuidae) | 1 | - | 1 | - | - | - | - | - | - | Нет |
| Кархаринообразные (Сarcharhiniformes) | ||||||||||
| Кошачьи акулы (Scyliorhinidae) | 86 | 13 | 45 | 4 | - | - | - | - | - | Эпибенталь |
| Просциллиевые (Proscyllidae) | 6 | 3 | 1 | - | - | - | - | - | - | " |
| Ложнокуньи акулы (Pseudotriakidae) | 1 | - | 1 | - | - | - | - | - | - | Нет |
| Куньи акулы (Triakidae) | 34 | 5 | 1 | - | - | - | - | - | - | Эпибенталь |
| Гнюсообразные (Torpediniformes) | ||||||||||
| Гнюсовые скаты (Torpedinidae) | 15 | 5 | 1 | - | - | - | - | - | - | " |
| Нарковые скаты (Narkidae) | 25 | 6 | 2 | - | - | - | - | - | - | " |
| Скатообразные (Rajiformes) | ||||||||||
| Ромбовые скаты (Rajidao) | 200 | 70 | 65 | 15 | - | - | - | - | - | Эпибенталь |
| Ложноромбовые скаты (Pseudorajidae) | 4 | - | 4 | - | - | - | - | - | - | Нет |
| Нитерылые скаты (Anacanthohatidae) | 15 | 2 | 13 | - | - | - | - | - | - | " |
| Xвостоколообразные (Myliobatiformes) | ||||||||||
| Шестижаберные скаты (Hexatrygonidae) | 3 | - | 3 | - | - | - | - | - | - | Нет |
| Химераобразные (Chimaeriformes) | ||||||||||
| Химеровые (Chimaeridae) | 20 | 2 | 18 | - | - | - | - | - | - | " |
| Носатые химеры (Rhinochimaeridae) | 6 | - | 4 | 2 | - | - | - | - | - | " |
| Спиношипообразные (Notacanthiiormes) | ||||||||||
| Галозавровые (Halosauridae) | 15 | - | 4 | 9 | 1 | 1 | - | - | - | " |
| Спиношиповые (Notacanthidae) | 9 | 2 | 3 | 4 | - | - | - | - | - | " |
| Липогеновые (Lipogenydae) | 1 | - | 1 | - | - | - | - | - | - | " |
| Угреобразные ( Anguilliformes) | Слитножаберные угри (Synaphobranchidae) | 28 | 2 | 7 | 4 | 1 | 6 | - | - | - | Эпибенталь |
| Неттастомовые (Nettastomatidae) | 35 | 4 | 12 | 4 | - | - | - | - | - | " |
| Конгеровые (Congridae) | 110 | 20 | 20 | - | - | - | - | - | - | " |
| Моногнатовые (Monognathidae) | 14 | - | 3 | 2 | 4 | 2 | 3 | - | - | Нет |
| Лососеобразные (Salmoniformes) | ||||||||||
| Серебрянковые (Argentinidae) | 35 | 15 | 7 | - | - | - | - | - | - | Мезопелагиаль |
| Гладкоголововые (Alepocephalidae) | 98 | 3 | 40 | 30 | 20 | 5 | - | - | - | Нет |
| Лептохиловые (Leptochilichthyidae) | 2 | - | 1 | - | - | 1 | - | - | - | " |
| Платитроктовые (Platytroctidae) | 40 | 1 | 27 | 10 | 2 | - | - | - | - | " |
| Стомиеобразные (Stomiiformes) | ||||||||||
| Гоностомовые (Gonostomatidae) | 26 | - | 3 | - | - | - | - | - | - | Мезобатипелагиаль |
| Топориковые (Sternoptychidae) | 51 | - | 37 | - | - | - | - | - | - | Мезобатипелагиаль |
| Фотихтовые (Photichthyidae) | 21 | - | 4 | - | - | - | - | - | - | " |
| Астронестовые (Astronesthidae) | ||||||||||
| Меланостомиевые (Melanostomiatidae) | 27 | - | 5 | - | - | - | - | - | - | Мезопелагиаль |
| Ayлапообразные (Aulopiformes) | ||||||||||
| Аулоповые (Aulopidae) | 10 | 5 | 1 | - | - | - | - | - | - | Эпибенталь |
| Зеленоглазковые (Chlorophthalmidae) | 45 | 20 | 7 | - | - | - | - | - | - | Нет |
| Нотосудиевые (Notosudidae) | 19 | - | 17 | - | - | - | - | - | - | Мезопелагиаль |
| Ящероголововые (Synodontidae) | 40 | 6 | 1 | - | - | - | - | - | - | Эпибенталь |
| Миктофообразные (Myctophiformes) | ||||||||||
| Неоскопеловые (Neoscopelidae) | 5 | - | 4 | - | - | - | - | - | - | Мезопелагиаль |
| Миктофовые (Myctophidae) | 220 | 8 | 12 | - | - | - | - | - | - | Мезобатипелагиаль |
| Трескообразные (Gadiformes) | ||||||||||
| Паркетниковые (Muraenolepididae) | 4 | 1 | 1 | - | - | - | - | - | - | Эпибенталь |
| Моровые (Moridae) | 90 | 20 | 40 | 15 | - | - | - | - | - | " |
| Меланоновые (Melanonidae) | 2 | - | 1 | - | - | - | - | - | - | Мезопелагиаль |
| Брегмацеровые (Bregmacerotidae) | 7 | - | 5 | - | - | - | - | - | - | " |
| Тресковые (Gadidae) | 50 | 12 | 3 | - | - | - | - | - | - | Эпибенталь |
| Мерлузовые (Merlucciidae) | 25 | 24 | 1 | - | - | - | - | - | - | Нет |
| Долгохвостовые (Macrouridae) | 350 | 10 | 245 | 45 | 15 | 30 | 5 | - | - | " |
| Ошибнеобразные (Ophldiiformes) | ||||||||||
| Ошибневые (Ophidiidae) | 170 | 15 | 35 | 30 | 20 | 30 | 12 | 3 | - | Эпибенталь, мезопелагиаль |
| Бититовые (Bythitidae) | 85 | 13 | 13 | 3 | 2 | 2 | - | - | - | Эпибенталь |
| Афионовые (Aphyonidae) | 18 | - | 5 | - | 2 | 3 | 8 | - | - | Нет |
| Удильщикообразные (Lophiiformes) | ||||||||||
| Удильщиковые (Lopliiidae) | ||||||||||
| Удильщиковые (Lopliiidae) | 25 | 12 | 8 | 1 | - | - | - | - | - | Эпибенталь |
| Хаунаксовые (Chaunacidae) | 10 | 2 | 8 | - | - | - | - | - | - | Нет |
| Нетопыревые (Ogcocephalidae) | 60 | 12 | 26 | 7 | - | - | 1 | - | - | Эпибенталь |
| Тауматихтовые (Thaumatichthyidae) | 5 | - | - | - | - | - | 1 | - | - | Мезобатипелагиаль |
| Дицерациевые (Diceratiidae) | 5 | - | - | 5 | - | - | - | - | - | Нет |
| Китовидкообразные (Cetomimiformes) | ||||||||||
| Ателеоповые (Ateleopidae) | 12 | 3 | 9 | - | - | - | - | - | - | " |
| Бериксообразные (Berycidae) | ||||||||||
| Трахихтовые (Trachichthyidae) | 38 | 11 | 21 | - | - | - | - | - | - | Эпибенталь |
| Диретмовые (Diretmidae) | 4 | - | 1 | 2 | - | - | - | - | - | Мезопелагиаль |
| Бериксовые (Berycidae) | 8 | 7 | 1 | - | - | - | - | - | - | Нет |
| Стефанобериксовые (Stephanoberycidae) | 3 | - | - | 1 | - | 2 | - | - | - | " |
| Меламфаевые (Melamphaidae) | 33 | - | 2 | - | - | - | - | - | - | Мезабатипелагиаль |
| Гиспидобериксовые (Hispidoberycidae) | 1 | - | 1 | - | - | - | - | - | - | Нет |
| Гибберихтовые (Gibberichthyidae) | 2 | - | - | 2 | - | - | - | - | - | " |
| Солнечникообразные (Zeiformes) | ||||||||||
| Паразеновые (Parazentidae) | 1 | - | 1 | - | - | - | - | - | - | " |
| Зенионовые (Zeniontidae) | 4 | - | 4 | - | - | - | - | - | - | Нет |
| Циттомимовые (Cyttomimidae) | 2 | - | 2 | - | - | - | - | - | - | " |
| Лунниковые (Oreosomatidae) | 10 | - | 8 | 2 | - | - | - | - | - | " |
| Чешуеигловые (Grammicolepididae) | 4 | - | 4 | - | - | - | - | - | - | " |
| Капровые (Caproidae) | 11 | 9 | 2 | - | - | - | - | - | - | " |
| Скорпенообразные (Scorpaentformes) | ||||||||||
| Скорпеновые (Scorpaenidae) | 310 | 80 | 15 | 1 | - | - | - | - | - | Эпибенталь |
| Перистедиевые (Periostediontidae) | 25 | 10 | 10 | - | - | - | - | - | - | " |
| Плоскоголововые (Platycephalidae) | 60 | - | 1 | - | - | - | - | - | - | " |
| Аноплопомовые (Anoplopomatidae) | 2 | 2 | - | 1 | - | - | - | - | - | Нет |
| Рогатковые (Cottidae) | 300 | 45 | 10 | - | - | - | - | - | - | Эпибенталь |
| Психролютовые (Psychrolutidae) | 30 | 10 | 10 | 5 | - | - | - | - | - | " |
| Липаровые (Liparididae) | 150 | 25 | 60 | 25 | 12 | 5 | - | - | - | Эпибенталь |
| Окунеобразные (Perciformes) | ||||||||||
| Эпигоновые (Epigonidae) | 26 | 3 | 22 | 1 | - | - | - | - | - | Нет |
| Морские лещи (Bramidae) | 17 | - | 2 | - | - | - | - | - | - | Эпипелагиаль |
| Лжесельдевые (Bathyclupeidae) | 4 | - | 3 | 1 | - | - | - | - | - | Нет |
| Щетинозубовые (Chaetodontidae) | 114 | - | 1 | - | - | - | - | - | - | Эпибенталь |
| Кабан-рыбы (Pentacerotidae) | 12 | 5 | 3 | - | - | - | - | - | - | " |
| Цеполовые (Cepolidae) | 20 | 9 | 1 | - | - | - | - | - | - | " |
| Губановые (Labridae) | 500 | 5 | 1 | - | - | - | - | - | - | " |
| Бельдюговые (Zoarcidae) | 160 | 50 | 45 | 20 | 5 | 3 | 7 | - | - | " |
| Нототениевые (Nototheniidae) | 50 | 30 | 2 | - | - | - | - | - | - | " |
| Усатые дракончики | 15 | 8 | - | 1 | - | - | - | - | - | " |
| Батидраковые (Bathydraconidae) | 16 | 3 | 2 | 3 | 1 | - | - | - | - | Нет |
| Белокровные рыбы (Channichthyidae) | 16 | 6 | 4 | 4 | 1 | - | - | - | - | Эпибенталь |
| Перкофисовые (Percophidae) | 35 | 15 | 15 | - | - | - | - | - | - | " |
| Драконетковые (Draconettidae) | 10 | 3 | 7 | - | - | - | - | - | - | Нет |
| Гемпиловые (Gempylidae) | 22 | 3 | 13 | - | - | - | - | - | - | Эпимезопелагиаль |
| Сабли-рыбы (Trichiuridae) | 27 | 3 | 21 | - | - | - | - | - | - | Эпибенталь |
| Центролофовые (Centrolophidae) | 22 | 9 | 4 | - | - | - | - | - | - | Эпипелагиаль |
| Камбалообразные (Pleuronectiformes) | ||||||||||
| Ботусовые (Bothidae) | 210 | 20 | 15 | - | - | - | - | - | - | Эпибенталь |
| Камбаловые (Pleuronectidae) | 100 | 49 | 12 | - | - | - | - | - | - | Эпибенталь |
| Циноглоссовые (Cynoglossidae) | 100 | 30 | 20 | - | - | - | - | - | - | " |
| Солевые (Soleidae) | 120 | 15 | 1 | - | - | - | - | - | - | " |
| Скалозубообразные (Tetraodontiformes) | ||||||||||
| Холлардиевые (Triacanthodidae) | 20 | 10 | 10 | - | - | - | - | - | - | Нет |
(Примечание. В таблицу не включены отряды и семейства, представленные только в переходной от шельфа к склону зоне (ЭМ) — миногообразные (Petromyzontiforrnes): миноговые (Petrornyzontidae) — 1 вид; многожаберникообразные: многожаберные акулы (Hexanchidae) — 4; катранообразные: звездчатошипые акулы (Echinorhinidae) — 2; плоскотелообразные (Spuatini-formes): плоскотелые акулы (Squatinidae) — 2; кархаринообразные: серые акулы (Carcharhinidae) — 1; скатообразные: новозеландские скаты (Arhynchobatidae) — 1: целакантообразные (Coelacanthiformes): латимерисвые (Latimeriidae) — 1; тарпонообразные (Elopjformes): гисовые (Pterothrissidae) — 2j угреобразные: ложнококгеровые (Xenocongridae) — 1, острохвостые угри (Ophichthidae) — 5. макроцефенхеловые (Macrocephenchelyidae) — 1: ошибнеобразные: карапусовые (Carapidae) — 5: удильщикообразные: клоуновые (Antennariidae) — 3; бериксообразаые: шишечниковые (Monocentridae) — 4, голоцентровые (Holocentridae) — 12, полимиксиевые (Polymixiidae) — 6; солнечникообразные: солнечниковые (Zeidae) — 10; колюшкообразные (Gasterosteiformes): бекасовые (Macrorhamphosidae) — 8; скорпенообразные: конгиоподовые (Congiopodidae) — 5, тригловые (Triglidae) — 15, гоплихто-вые (Hoplichthyidae) — 10, терпуговые (Hexagrammidae) — 2, эреуниевые (Ereuniidae) — 2, лисичковые (Agontdae) — 15; окунеобразные: каменные окуни (Serranidae) — 40, полиприоновые (Polyprionidae) — 3, каллантиевые (Callanthiidae) — 10, приакантовые (Priacanthidae) — 5, акропомовые (Acropomatidae) — 15, мицуевые (Scombropidae) — 4, малакантовые (Malacanthidae) — 20, ставридовые (Carangidae) — 6, красноглазковые (Emmelichthyidae) — 11, луциановые (Lutianidae) — 10, спаровые (Sparidae) — 15, центракантовые (Gentracanthldae) — 2, джакасовые (Cheilodactylidae) — 5, трубачевые (Latriobidae) — 1, стихеевые (Stichaeidae) — 10, зубатковые (Anarhichadidae) — 4, хампсодоновые (Champsodontidae) — 6, волосозубовые (Trichodontidae) — 2, звездочетовые (Uranoscopidae) — 5, мугилоидовые (Mugiloididae) — 5, пескарковые (Callyonimidae) — 15, бычковые (Gobiidae) — 10, ариоммовые (Ariommidae) — 10; камбалообразные: безрукие камбалы (Achiropsettidae) — 12; скалозубообразные: триодоновые (Triodontidae)— 1. Эти 345 видов включены в итоговую строку колонки ЭМ.)
Изучение рыб подводных поднятий только начинается, но уже сейчас можно отметить, что в разных районах Мирового океана в заселении этого биотопа приняли участие в основном представители одних и тех же, притом сравнительно немногих, семейств. Как показывает сравнение ихтиофаунистических списков подводных гор Метеор и Жозефина в северо-восточной Атлантике, банки Вальдивия Китового хребта в юго-восточной Атлантике, тихоокеанских подводных хребтов Кюсю-Палау, Гавайского, Северо-Западного, Наска и Сала-и-Гомес, сходство в составе населения этих далеко отстоящих друг от друга участков довольно велико, особенно на уровне семейств и родов.
Видовой состав ихтиоценов на подводных поднятиях, определяется в первую очередь глубиной над их вершинами, а разнообразие этого состава — действием «островного эффекта», прямо зависящего от площади поднятия Талассоэпибентальные ихтиоцены на вершинах самых мелководных банок (например, гор Келсо, Кейпл и Уанганелла в Тасмановом море, имеющих глубину 40—100 м) обычно представляют собой в той или иной степени обедненный вариант сообществ ближайших шельфовых районов. Не исключена возможность, что на крохотных вершинных площадках постоянные самовоспроизводящиеся популяции эпибентальных рыб вообще отсутствуют и их население пополняется только за счет оседания пелагических личинок, принесенных извне, т. е. с ближайшего шельфа.
Существование независимых горных популяций рыб эпимезо- и мезобентопелагического комплекса, напротив, не вызывает никаких сомнений. Об этом свидетельствуют не только многолетние наблюдения над локальными популяциями таких, к примеру, видов, как рыба-кабан Pseudopentaceros wheeleri на Гавайском подводном хребте, капродон Caprodon longimanus на уже упоминавшейся банке Уанганелла, безпузырный окунь Helicolenus lengerichi и зеркальный солнечник Zenopsis sp. (неописанный вид, близкий к Z. nebulosus) на подводном хребте Наска, но и существование эндемичных видов, ареалы которых ограничены одним поднятием или группой близкорасположенных поднятий. Эндемичные талассобатиальные виды известны сейчас из многих районов Мирового океана, в том числе с гор Азорского архипелага и с Китового хребта в Атлантическом океане, Мадагаскарского и Восточно-Индийского хребтов в Индийском океане, хребтов Кюсю-Палау, Северо-Западного, Наска и Сала-и-Гомес в Тихом океане. Последний район — хребты тепловодной юго-восточной Пацифики — представляется, кстати сказать, прекрасным местом для разнопланового модельного изучения талассных ихтиоценов, в том числе и для ответа на вопросы, поставленные К. Хаббзом. Уникальность этих хребтов состоит в том, что, несмотря на близость к материку Южной Америки, они заселены рыбами индовестпацифического происхождения, и поэтому ближайшие шельфы и склоны явно не поставляют никакого материала для пополнения их населения. Сами эти горы представляют собой погруженные коралловые острова (гайоты), различающиеся по глубине и возрасту (геологическому времени существования).
Весной 1980 г. в нашу лабораторию поступили материалы, несколькими месяцами раньше собранные моими бывшими аспирантами Г. А. Голованем (теперь он возглавляет научные работы в Севастопольском бюро по подводным исследованиям) и Н. П. Пахоруковым. Они принимали участие в рейсе «Ихтиандра» на подводные хребты Наска и Сала-и-Гомес, которые до того были совсем неизученными в ихтиологическом отношении (правда, еще раньше там работали сотрудники ТИНРО, но, к большому сожалению, они, как и во многих других рыбохозяйственных экспедициях, не собирали представительных коллекций рыб). Предварительная обработка полученных нами проб (ее проводили Ю. Сазонов, Ю. Щербачев и я) дала на редкость интересные и, я бы сказал, неожиданные результаты. Выяснилось, что среди донных и придонных рыб этого района восточной Пацифики преобладают виды, которые были известны только из западной и центральное частей океана — от Японии, Австралии, Гавайских островов, Новой Каледонии, а многие другие относятся к числу неописанных; Впоследствии состоялось еще несколько экспедиций в этот рая он - я сам побывал там дважды на «Профессоре Месяцеве» и «Профессоре Штокмаые» и в результате почти полностью переключился на изучение «горных» рыб. Сейчас список видов, обнаружейных на обоих хребтах, увеличился до 160, и почти треть из них — это исключительно высокий процент — оказалась эндемичной (описания 30 новых видов уже опубликованы, остальные готовятся к печати). В моих ближайших планах одно из первых мест занимает сейчас подготовка монографии «Рыбы подводных хребтов Наска и Сала-и-Гомес», над которой мы работаем вмести с сотрудником ВНИРО А. Н. Котляром (у него я тоже был когда-то руководителем по кандидатской диссертации), надеясь завершить ее через год — другой.
И еще: не могу не упомянуть о трудностях тралений н гоpax. Представление об этих трудностях легко можно получить вообразив, что нужно провести трал с дирижабля, летящего выше сплошного облачного покрова, по склонам, скажем, Кавказского хребта. Эта мысль постоянно приходит мне в голову, когда я смотрю на горы в иллюминатор самолета, и такое действие представляется просто невероятным, но тем не менее... При первых в моей практике работах на подводных поднятиях (это был горный массив Экватор в западной части Индийского океана) мы раз за разом опускали с «Рифта» оттертрал на склоны (естественно, не вслепую, а ориентируясь по эхолоту), порвали все, что рвется но так и не смогли получить полноценной пробы. Особенно запомнился один задев, когда траловая доска, попавшая в расщелину, превратилась у нас в мертвый якорь и вышедшее на верти кальный трос (моряки говорят «на панер») и содрогающееся oт напряжения судно не могло на любом курсе сдвинуться с местаЛ пока 25-миллиметровый стальной ваер не лопнул с таким громоподобным звуком, что создалось полное впечатление взрыва. Хорошо работать только на ровных вершинах гайотов, да и там трал могут подстерегать разнообразные и неожиданные сюрпризы Среди рыб, составляющих постоянное население вершин подводных гор, есть немало чисто донных видов (в том числе зеленоглазки, хаунаксы, скорпениды, морские петухи, камбалы), но наиболее массовый и характерпый компонент сообщества составляют бентопелагические объекты — колючие акулы, мавролик, специфичные для гор рыбы-топорики и светящиеся анчоусы, рыбы-бекасы мориды, долгохвосты, солнечники, бериксы, большеголовы, некоторые ставридовые, каменные окуни, эпигонусы, красноглазки, рыбы-сабли, масляные рыбы и др.
Биология большинства «горных» бентопелагических рыб пока изучена слабо. Известно, правда, что многие из них совершают суточные вертикальные миграции: днем они держатся вблизи дна, а в темное время поднимаются в верхнюю толщу вод, удаляясь на 100—200 м и более от твердой поверхности. Как уже указывалось, такие миграции предпринимают даже мелкие стайные виды (Maurolicus muelleri, Diaphus suborbitalis). В связи с этим возникает естественный вопрос, как удается этим рыбам, несмотря на наличие течений, иногда достаточно сильных, всю ночь оставаться над поднятием, с которым они теряют визуальный контакт, и точно опускаться на него на рассвете. Все объяснения этого феномена (ориентация по светилам, поляризованному свету, градиентам скорости и т. п.) чисто умозрительны и выглядят неубедительно.
Возможность массовых миграций талассобатиальных рыб от одного подводного поднятия к другому кажется мне весьма проблематичной, если не говорить о горах, расположенных на одном цоколе: в этом случае эврибатные виды (низкотелый берикс, например, обитает в диапазоне от 50 до 1000 м), естественно, могут перемещаться в пределах всего района, подходящего по глубине. Эта точка зрения не является, однако, общепринятой. Предполагают, в частности, что рыба-кабан Гавайского подводного хребта проводит от 4—5 до 7 лет в пелагиали северной Пацифики и только после этого приходит на горы, где размножается. Весьма логичная схема функциональной структуры попу-ляционных ареалов берикса в Атлантическом океане, в основу которой положены данные о размерно-возрастном составе и зрелости рыб в разных районах, была недавно разработана Ф. Е. Алексеевым с соавторами. Вот как описывается, например, цикл существования североатлантической популяции: нерестовая часть ареала приурочена к горам Углового поднятия в западной части круговорота; оттуда пелагические икра, личинки и мальки дрейфуют в Северо-Атлантическом течении на восток и юго-восток; молодь оседает на подводных поднятиях в восточной части круговорота, а в отдельные годы и на склоне северозападной Африки; по мере полового созревания берикс активно мигрирует на запад по южной периферии круго-ворота, и половозрелый контингент постоянно обитает в Репродуктивной области (т. е. на Угловом поднятии). Я думаю, что региональные различия в размерах берикса, взятого из промысловых уловов, могут объясняться разное глубиной лова (половозрелые особи обитают глубже, чем созревающие, а траловые участки на Угловом поднятии находятся на большей глубине, чем на Азорских банках) Большое сомнение вызывает и сама способность мигрантов к отысканию нерестовых участков — для этого нужно предположить почти сверхъестественные навигационные cпoсобности рыб. И наконец, этому представлению, как и всем основанным на косвенных доказательствах соображениям в пользу существования таких миграций, противоречит полное отсутствие этих рыб над большими глубинами в открытом океане. До сих пор не отмечено ни одной; поимки взрослых бериксов, красноглазок и подобных им объектов в далеком удалении от материковых склонов подводных возвышенностей.
Хуже всего изучены жизненные циклы бентопелагических рыб и особенно их размножение и ранние стадии онтогенеза, во многом загадочные в связи с исключительно редкой встречаемостью личинок и мальков в уловах, полученных любыми орудиями лова. Относительно воспроизводства рыб, обитающих на подводных поднятиях, высказывались два предположения. Согласно одной точке зрения, развитие их планктонных стадий происходит системах местных циркуляции вблизи гор. Альтернативная гипотеза о заселении поднятий личинками и мальками, приносимыми течениями «извне», также имеет своих сторонников.
Действительно, слабая изученность процессов воспроизводства талассобентальных рыб сильно препятствует однозначному решению вопроса. В то же время доказанпость существования независимых популяций на отдельных банках, например на горе Метеор в Атлантическом; океане или на некоторых группах близкорасположенных поднятий хребтов Наска и Сала-и-Гомес, дает полное ocнование отказаться от представления об основополагающей роли разноса икры, личинок и мальков течениями и наличии значительного обмена генофондами. Развития планктонных стадий может, по-видимому, происходите в непосредственной близости от подводных возвышенностей в локальных круговоротах любого пространственного масштаба, даже не имеющих стационарного характера, но обладающих такой временной устойчивостью, которая была бы соизмерима с продолжительностью пассивного пребывания икры и личинок в пелагиали (около 10—15 суток). Еще одна гипотетическая возможность, требующая экспериментальной проверки, состоит в откладывании икринок с отрицательной плавучестью в щели и углубления дна. Более того, как показало исследование разработанной В. Б. Цейтлиным и Ю. А. Рудяковым имитационной модели независимой популяции рыб, населяющих подводную гору, такая популяция вполне способна сохраняться и воспроизводиться даже и при разносе рыбной молоди на планктонных стадиях развития течениями, если благодаря турбулентной диффузии она будет иметь шанс случайно вернуться к месту рождения. Из этой модели следует, правда, что вниз по течению от горы должен вытягиваться довольно значительный «шлейф» икры и личинок, чего в природе никогда не наблюдали.
Итак, что же прояснилось в наших знаниях о рыбах подводных поднятий за три десятка лет, прошедших с того времени, когда Карл Хаббз задал свои вопросы? Стало ясно, что видовой состав ихтиоценов отдельных поднятий характеризуется значительным постоянством во времени, ибо каждая гора представляет собой особый биотоп с населяющим его биоценозом. Заселение гор происходило точно тем же путем, что и заселение островов и их цоколей — с шельфов и склонов материков, и поэтому подводные поднятия должны рассматриваться в том же ряду природных объектов, что и океанические острова. Колонизация островных биотопов в открытом океане (включая горы) шла в основном за счет более разнообразных прибрежных фаун, населяющих западные части океанов, и чем дальше расположено поднятие от главного источника своего формирования, тем выше эндемизм его населения (максимальная доля эндемиков — около 40% в диапазоне глубин 200—500 м выявлена на хребтах Наска и Сала-и-Гомес). Поднятия, конечно же, являются «опорными пунктами» в расселении тех рыб, которые способны к достаточно долгому выживанию в пелагиали во взрослом состоянии или на стадии молоди (хороший пример дают поимки в пелагиали восточной тропической Пацифики крупных мальков большеглазой каталуфы Cookeolus boops, явно происходящих от популяций подводных гор). Также очевидно, что среди среднеглубинных видов есть потомки мелководных форм (уже упоминавшийся щетинозуб Chaetodon guyotensis с хребта Кюсю-Палау — типичный дериват фауны коралловых рифов), возникшие в результате заглубления вершин поднятий. Не вызывает никаких сомнений и вполне очевидный сейчас факт видовой дивергенции рыб в условиях изоляции на горах, имеющих достаточно длительную историю существования. Вопрос о скоплениях рыб у подводных поднятий тоже не остался без ответа (этот аспект обсуждается в гл. VI). Таким образом, все «загадки» К. Хаббза вроде бы разгаданы, но полной ясности в талассобатиальной проблеме все еще нет: на передний план выдвинулись теперь экологические aспекты существования горных популяций. Будущие ихтиологические изыскания на океанских «банках» должны быть направлены прежде всего на изучение размножения, жизненных циклов, миграций и особенностей поведения рыб (здесь никак нельзя обойтись без использования обитаемых подводных аппаратов), а также на выявление популяционной структуры отдельных видов.
Биотопическое своеобразие подводных поднятий, о котором шла речь выше, не может затушевать, однако, очевидного сходства их населения с населением материковых склонов в тех же диапазонах глубин. Повсюду самая верхняя часть среднеглубинной зоны имеет переходный (иначе говоря, экотонный) характер, по батиметрические границы экотона и степень его своеобразия, которую можно выразить процентом видового эндемизма в пределах эпимезобентопелагического комплекса рыб, насчитывающего более 1100 видов (см. табл. 4), сильно варьируют в разных участках Мирового океана.
Ихтиоцены талассной мезобентали отличаются, насколько можно судить по имеющимся данным, главным образом заметным видовым обеднением по сравнению с прилегающими участками континентальных склонов. На Китовом хребте, например, на глубине 200—1500 м отмечено всего 60—70 видов рыб, что в 3—4 раза меньше, чем на соответствующих глубинах склона юго-западной Африки. По Ю. Н. Щербачеву, из 36 видов батиальных офидиид Индийского океана 20 найдены пока только на материковом склоне и лишь 3 только в талассобатиали (13 видов являются общими). Это обеднение связано не только с отсутствием видов, проникающих в мезобеиталь с шельфа, но, по-видимому, отражает и меньшую «емкость» талассобатиали — меньшее разнообразие экологических ниш.
Мезобенталь и прилегающая к ней водная толща заселены хорошо обособленным сообществом рыб, в состав которого входит почти 1200 специфических видов. Правда, число эндемичных для этого биотопа семейств невелико, по к нему приурочено наибольшее разнообразие многих групп. Особенно характерны для этой зоны колючие акулы, в частности из рода Etmcpterus, кошачьи акулы Apristurus, ромбовые скаты, слитиожаберыые угри, гладкоголовы, моровые, долгохвосты (наиболее богатая видами группа донно-придоиных глубоководных рыб), ошибневые, бериксообразпые, солнечиикообразные и др.
Большая часть рыб, населяющих верхнюю часть склона на глубине менее 1500 м, принадлежит к придонным и донным видам, однако среди них есть, как уже говорилось, и чисто пелагические формы, не имеющие непосредственной связи с дном и в какой-то степени аналогичные неритическим рыбам эшгаелагиали. Помимо настоящих пектонных рыб — колючих акул, долгохвлстов, лунников, гемпиловых и др., к этой мезобентопелагической группировке относятся и макропланктонные виды, близкородственные видам, характерным для океанской мезопелагиали. В их число входят, напомню, представители таких семейств, как топориковые (в том числе мавролик), фотихтовые, меланостомиевые, миктофовые, меламфаевые, меланоновые и др. Предполагалось, что основы ареалов этих макропланктонных видов находятся в системе горизонтальных и вертикальных вихрей, существующих над материковым склоном, но прямых доказательств этому нет. Отсутствие у большинства таких рыб значительного выноса за пределы биотопа (вод, покрывающих склоны и подводные поднятия) на любых стадиях жизненного цикла свидетельствует о существовании специального механизма удержания их в присклоновой зоне, сущность которого остается пока непонятной.
Общее число рыб, обитающих в мезобеитали и мезобентопелагиали, намного больше, чем в других океанических ихтиоцеиах. По довольно приблизительному (но отнюдь не завышенному) подсчету (см. табл. 4), в этой зоне обитают около 2600 видов (почти 1500 без экотонпых шелъфо-склоновых рыб). Обособленность этой фауны сильнее всего выражена, на видовом уровне, хотя число родов, обнаруженных только в водах верхней части материкового склона и подводных поднятий, также достаточно велико. В то же время к числу специфических мезобентальных таксонов более высокого уровня можно отнести только относительно немногие семейства, в то время как остальные группы представлены либо па шельфе, либо в нижней части склона, а часто и в обоих биотопах.
В сообществах нижней части склона (батибенталь) и над ней (батибентопелагиаль) представлены сравнительно немногие систематические группы. Из хрящевых рыб здесь живут только колючие акулы из родов Centropho rus, Centroscymnus, Somniosus, кошачьи акулы Apristurus, ромбовые скаты Bathyraja и Breviraja, а также носатые химеры-харриотты, по ни один вид не является эндемичным для биотопа. Среди костистых рыб наиболее характерны виды родов Aldrovandia, Synaphobranchus, Alepocephalus, Bathypterois, Bathysaurus, Antimora, Coryphaenoides, Cataetyx, Paraliparis, Lycenchelys и др. По числу видов и особей в тропиках явно доминируют три группы — гладкоголовы, долгохвосты и ошибневые, а в умеренных и холодных водах две — морские слизни и бельдюговые. Суммарное число видов достигает 450, но большая их часть (273 вида) оказывается общей с нижней мезобенталью (граница двух сообществ батиали представляет собой экотонную зону, расположенную на глубине 1300—1800 м), а многие другие (91 вид) — с абиссобенталью, так что на долю специфической фауны приходится менее 20% состава. В нижней части склона различия между континентальной и талассной батибентальпыми группировками еще слабее, чем в мезобентали. В Индийском океане, например, по данным Ю. И. Сазонова, около 75% видов гладкоголовов встречается как на склоне континентов, так и на подводных поднятиях.
Абиссобентальные и абиссобептопелагическяе ихтиоцены включают около 140 видов, однако эндемичные для ложа океана виды очень немногочисленны. Число специфических родов 10—12, среди них Ericara из гладкоголо-вых, Bathymicrops пз зеленоглазковых, Echinomacrurus пз долгохвостовых, Acanthonus и Typhlonns из ошибневых, Pachycara и Thermarces из бельдтоговых. Остальные виды принадлежат к родам, представленным и в батибептали (Synaphobranchus, Einoctes, Bathytyphlops, CoryI haenoides).
При изучении сообществ термальных источников, обнаруженных недавно в Тихом и Атлантическом океанах па глубине более 2000 м, выяснилось, что в состав этих биоценозов входят и рыбы. Как показали подводные наблюдения, фотосъемки и ловы, вблизи геотермальных излияний Галапагосского рифта встречено более 20 видов рыб (в основном макрурид и офидиид), но почти все они держатся в некотором удалении от мест вытекания теплой, насыщенной сульфидами и лишенной кислорода воды. Единственное исключение составляли особи одного вида, предварительно отнесенные к роду Diplacanthopoma (пока не удалось поймать ни одного экземпляра), которые по-одиночке, парами и тройками постоянно появлялись из кратера источника и снова уходили внутрь. Предполагается, что эта рыба питается непосредственно бактериями, окисляющими сероводород. В термальной воде живет также зоарцида Thermarces cerberus, являющаяся, по всей вероятности, постоянным членом этого своеобразного сообщества; другой представитель того же рода обнаружен только в периферической зоне рифтового «оазиса». Слабовыраженный (только на родовом и видовом уровне) эндемизм гидротермальных рыб резко контрастирует с большой обособленностью бентосных беспозвоночных (подкласс погонофор, надсемейство моллюсков и т. д.).
В глубоководных желобах (хадобенталь) до сих пор обнаружено всего 7 видов рыб, ведущих дойный образ жизни (рис. 42). В их числе три вида ошибневых — Leucicorus atlanticus (наибольшая глубина вылова 6800 м в желобе Пуэрто-Рико), Holcomycteronus profundissimus (7160 м в желобе Сунда у южного побережья острова Ява) и Abyssobrotula galatheae (8370 м в желобе Пуэрто-Рико) и четыре вида морских слизней — Notoliparis macquariensis из желоба Маккуори (глубина 5410 м), N. kurchatovi пз Южно-Сандвичева желоба (5470 м) (в табл. 4 эти два вида условно помещены в графу АХ), N. kermadecensis из желоба Кермадек (6770 м) и Pseudoliparis amblysto-mopsis и неописанный вид этого же рода из Курило-Кам-чатского и Японского желобов (максимальная глубина лова почти 7580 м). Все хадобентальные офидииды встречаются и в абиссобентали (рекордист по глубине — абиссобротула — добывалась в восточной Пацифике всего лишь на глубине 2330 м), тогда как оба рода липарид (они, кстати сказать, описаны А. П. Андрияшевым) эндемичны для глубин более 5500—6000 м. Насколько сейчас известно, основную пищу всех этих видов составляют донные ракообразные (изоподы и амфиподы), но у кермадекских нотолипарисов, а их было поймано сразу 5 в одном трале, в желудках оказались также рыбья чешуя и кости. Хищничество представляется, однако, очень маловероятным для этих некрупных и к тому же вялых и пассивных, судя по всем данным, существ. Их поймали скорее всего там, где они собрались для поедания падали, опустившейся из верхних слоев океана. В яичниках двух самок этого вида было обнаружено 737 и 849 икринок, из которых самые крупные имели 7,5—9 мм в диаметре. И. Нильсен предполагал, что этот вид, подобно батиабиссобентальному карепрокту Careproctus ovigerum, может вынашивать развивающуюся икру в ротовой полости. Можно думать, что в хадобентали существуют в другие виды рыб, но их число вряд ли может быть значительным.
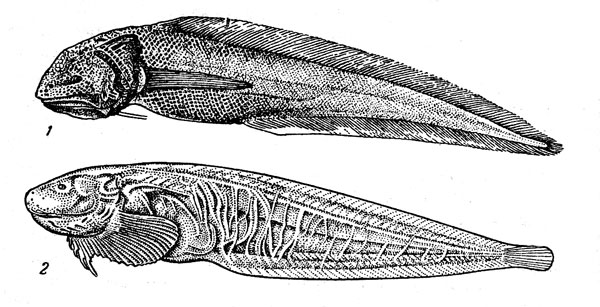
Рис. 42. Рыбы наибольших глубин абиссобротула (1) и псевдолипарис (2).
Все ловы донных рыб в глубоководных желобах выполнены с помощью очень примитивною и, казалось бы, анахроничного орудия лова, которое и в наш век технического прогресса остается таким же, как в прошлом столетии. Это — трал Сигсби, он же трал Агассица, названный так по фамилиям капитана американского корабля «Альбатрос» и выдающегося зоолога, неоднократного руководителя экспедиций на этом же судне. Трал этот предназначен для сбора бентоса.
Представьте себе две крутых дуги из железных пластин, скрепленных трубами и тросами, к которым прикреплен короткий мешок из грубой дели (внутри него помещается мелкоячейная вставка). Эта снасть опускается па дно на длинном тросе и, будучи совершенно симметричной (у нее нет ни верха, ни низа), всегда занимает на грунте рабочее положение. Само траление носит пассивный характер: судно, дрейфующее по ветру, медленно волочит трал, собирающий со дна все, что «плохо лежит» (точнее говоря, то, что лежит хорошо и не способно отплыть или отползти).
Каждое траление в ультраабиссали - большое событие в экспедиционной жизни. Посадка трала Сигсби па дно узкого желоба без применения специальной акустической аппаратуры (а она еще не нашла широкого применения в нашей практике) очень трудна. Она требует точного расчета дрейфа судна, а также оценки необходимой длины вытравливаемого троса (эта оценка, производимая «на глаз»» с учетом двух переменных - глубины по эхолоту и угла наклона троса, уходящего в воду,— самое слабое звено методики работ). Бывает, что трал не достигает дна или. наоборот, лишний трос завивается в кольца, «колышется» и рвется при
выборке, так что долгие часы работы пропадают впустую. Мне приходилось присутствовать при нескольких десятках удачных тралений в самых разных желобах (в том числе и в самом глубоком из них - Марианском), по только дважды - в Японском и Южно-Сандвичевом желобах они приносили рыб. Ясно, что на хадальных глубинах их совсем не так уж много.
Экология донно-придонных рыб открытого океана исследована, пожалуй, еще хуже, чем экология рыб глубоководной пелагиали. Данных о их размножении, росте, питании и других биологических особенностях пока немного, и общая картина, складывающаяся из разрозненных фрагментов, не может считаться вполне адекватной: многие вопросы еще ждут ответа. Тем пе менее уже сейчас выявлены некоторые общие закономерности, характерные для рыб бентопелагического комплекса.
Размножение. Хрящевые рыбы, обитающие в придонных слоях, демонстрируют все три возможных в этой группе типа репродукции — яйцерождение, яйцеживорождение и живорождение. К числу яйцеживородящих принадлежит п латимерия, но из костистых рыб этим путем размножаются только некоторые эпимезобентальные морские окуни (Sebastes) представители семейств Bythitidae и Aphyonidae. Преобладающее большинство видов имеет пелагические икринки, число которых сильно варьирует: от нескольких тысяч (у аргентин и батитифлопсов) до сотен тысяч (у моровых, долгохвостов, крупных камбаловых). У рыб, откладывающих донную икру (икру с отрицательной плавучестью), плодовитость, как правило, заметно меньше и измеряется десятками — сотнями икринок (минимально — 20 икринок у Rinoctes nasutns), по у некоторых видов гладкоголовов (крупные представители рода AlepocepLalus) может достигать 10 тыс.
Судя по встречаемости личинок и мальков в уловах, донно-придонных рыб можно разделить на две группы. У одних постэмбриональное развитие проходит в толще воды, у других — в придонных слоях. К первой группе, для которой характерны значительные морфологические преобразования в конце личиночно-малькового периода (метаморфоз), должны быть отнесены все угри и родственные им галозавриды и нотакантиды, также имеющие лептоцефальную стадию, некоторые хлорофтальмиды (Bathytyphlops с мальком макристиеллой) и сиподонтиды (Balhysaurus с мальком макристиумом), те офидииды, которые обладают специфическими личиночными стадиями типа «эксериллиум» или «вексиллифер», а также, по-видимому, мориды и долгохвосты. Вторую группу образуют алепоцефалиды, хлорофтальмиды из родов Ipnops и Bathymicrops, некоторые офидииды, все липариды и зоарциды и др.
Эта простая классификация не может быть распространена, однако, на всех рыб бентопелагического комплекса, поскольку у многих из них ранние стадии развития все еще остаются неизвестными. Она не является бесспорной даже в отношении некоторых из перечисленных выше рыб. Представлениям о наличии пелагической стадии у всех видов семейств Moridae и Macrouridae противоречит, например, существование в этих группах локального эндемизма: в обоих семействах сейчас известно немало видов, которые ограничены в своем распространении препятствиями, непреодолимыми для них, казалось бы, только во взрослом состоянии — для батибентальных рыб это мелководные пороги, отделяющие глубинные бассейны, для мезобентальных — океанические глубины, окружающие подводные возвышенности. Личиночно-мальковые стадии всех массовых доино-придопных рыб, особенно населяющих батиаль, вообще очень редки в толще воды и почти никогда не встречаются вдали от материковых склонов и, как уже говорилось, горных поднятий в открытых водах.
В связи с этим высказывались предположения о том, что эмбриональное и раннее постэмбриональное развитие таких рыб может проходить в придонных горизонтах.
Размеры и рост. В среднем донно-придонные глубоководные виды рыб крупнее пелагических глубоководных, но мельче тех, которые живут в верхней толще воды (см. рис. 27). Среди них нет таких гигантов, как эпипелагические акулы, марлины и меч-рыбы, но некоторые из батиальных видов достигают все же значительной длины. К числу наиболее крупных относятся полярная акула (длина 6,4 м при массе около 1 т), шестижаберная акула Нехаnchus griseus (4,8 м), акула-домовой (3,3 м), ложнокунья акула (3 м), звездчатошипая акула Echinorhinus brucus (2 м), ромбовые скаты рода Bathyraja (2 м), химеры 1(1,5 м), а также латимерия (длина 2 м, масса почти 100 кг). Из костистых рыб наибольших размеров достигают белокорый палтус (длина до 4,7 м и масса более 300 кг), клыкач (2,1 м), некоторые долгохвосты, ателеоиовые и гемпилиды (по 2 м), слитиожаберные угри (1,8 м), зубатки (1,4 м), иекоторые офпдииды (до 1,3 м), а самыми маленькими являются бентопелагические стерпоптихиды и миктофиды из родов Maurolicus, Polyipmis, Diaphus, Laitipanyciodos, а также бептические липариды и перцихтиды из родов Paraliparis, Osopsaron, Pteropsaron, длина которых не превышает 5—6 см. В абиссали размерный спектр значительно уже: от 19—20 см у некоторых гладкоголовов (Rmoctes, Mirognathus) до 85 см (при массе 7 кг) у батизавра Bathysaurus mollis, который, как показано прямыми наблюдениями с подводного аппарата, способен к резким рывкам на расстояние 10—15 м и, без сомнения, должен быть отнесен к нектону (в толще воды нектонные рыбы отсутствуют в батипелагиали и ниже).
Глубоководные донные и придонные рыбы, как правило, отличаются медленным ростом и большой продолжительностью жизни. Исключение составляют, пожалуй, только мелкие беитопелагические виды (мавролик, светящиеся анчоусы) из семейств, характерных для открытой пелагиали, которые совершают суточные вертикальные миграции из придонных горизонтов в эпипелагиаль. Все они, подобно их чисто пелагическим родичам, созревают в возрасте, близком к годовалому, и живут ненамного больше. У таких рыб, как колючие акулы, аргентины, гладкоголовы, моровые, крупные макруриды (род Coryphaenoides), большеголовы, скорпеновые, камбалы, продолжительность жизни варьирует, насколько сейчас известно, от 10 до 30 и даже 40 лет, причем годовые приросты длины редко превышают 3—5 см. Половозрелыми опи становятся в возрасте 5—10 лет, а иногда я позже. Эти рыбы с полным правом могут быть отнесены к категории длинноцикловых.
Питание. Представляется естественным, что рыбы, живущие на дне или у дна, должны питаться в основном пищей, находящейся на грунте, т. е. бентосными организмами. Это вполне логичное заключение, которое, как правило, оправдывается в приложении к донпо-придонным рыбам шельфовой зоны, оказывается, однако, совсем не бесспорным, когда речь идет о глубоководных видах — почти все массовые формы потребляют в пищу пелагических животных (мезо- и макропланктон, микронектон). Такая «диета» характерна не только для рыб, поднимающихся в водную толщу, но и для многих постоянных обитателей дна.
Значительная часть рыб бентопелагического комплекса относится к числу хищников, поедающих крупную добычу — других рыб, головоногих, ракообразных. Наиболее глубоководными из донных хищников являются батизавры, в пище которых зарегистрированы слитножабериые угри, макруриды, лиценхелы, креветки, кальмары. Известен и способ их охоты: рыбы поджидают добычу, полустоя или лежа на дне в слегка наклонном положении (передняя часть тела приподнята), и ловят ее неожиданным стремительным броском, сопровождаемым хватательным движением челюстей. Некоторые хищники мезобентопелагиалн способны к совместному добыванию добычи. Так поступают, к примеру, некоторые колючие акулы, а также рексии. Эти рыбы режут крупную рыбу на куски своими мощными зубами, а затем заглатывают по частям, и части одной жертвы иногда обнаруживаются в желудках разных хищников, пойманных одним тралом.
Конечно, на тех участках дна, где имеется достаточное количество кормового бентоса, он также идет в пищу рыбам. Многие из преимущественно планктоядных видов, в частности гладкоголовы, долгохвосты, мориды, нередко разнообразят свое питание за счет донных моллюсков, полихет, ракообразных. Как показал Р. Хэдрич, отдельные группы видов (подроды) рода Coryphaenoides, различающиеся размерами и строением зубного аппарата, различаются и по характеру пищи. Мелкие виды со слаборазвитым зубным вооружением (например, С. carapinus) поедают главным образом бентос, в первую очередь офиур, тогда как более крупные виды с сильными зубами — пелагических ракообразных, рыб, головоногих. Преимущественно бентоядными являются многие офидииды, угри, камбалы, чисто бентоядными — галозавры, спиношипы, линариды, встречающиеся в связи с этим только на мягких грунтах.
Интересно сопоставить трофическую структуру глубоководного донного ихтиоцена и таксоцена беспозвоночных того же сообщества. Большая часть глубоководного макробентоса, по М. Н. Соколовой, входит в состав пищевой группировки детритоедов, питающихся органическим веществом донных осадков. Также довольно многочисленны среди донных беспозвоночных сестоноеды, пищу которых составляют взвешенные в воде органические частицы, включая живой планктон. Плотоядные беспозвоночные, потребляющие живую добычу и трупы, представляют меньшинство и в сессильвом (сидячем) и в вагильном бентосе, но решительно преобладают в нектобентосе, (это в основном придонные ракообразные). Среди рыб чистых детритоедов, по-видимому, нет, хотя остатки грунта, заглоченные, можно думать, вместе с другой пищей, не раз обнаруживались в желудках бентофагов (например, у спиношипов и галозавров). К числу сестонофагов-хватателей (почти что сессильных) можно отнести батиптеров, возвышающихся над дном на треножнике, составленном лучами хвостового и брюшных плавников, а возможно, и других глубоководных хлорофтальмид (кроме ипнопсов). Преобладают в ихтиоцене, конечно, плотоядные, среди которых следует выделять потребителей донных беспозвоночных, всеядных хищнп-ков-падалыциков и настоящих хищников, охотящихся только на крупную живую добычу.
Вершинные участки подводных поднятий, как правило, имеют скальное дно, полностью лишенное рыхлых осадков, что само по себе определяет доминирование рыб-планктофагов в сообществах таких биотопов. До самого недавнего времени бытовали два взгляда на источник пищевых ресурсов, определяющих существование и продукцию «горных» популяций рыб. Согласно одному из них, продуктивность поднятий определяется локальным орографическим эффектом — подъемом глубинных вод, обусловленным завихрением набегающего на гору потока постоянного течения и приводящим в действие механизм биологической продуктивности в его полном цикле: от повышения концентрации биогенных солей в эвфотическом слое к увеличению первичной продукции водорослей и за ее счет к увеличению биомассы зоопланктона. Для более точного («более наукообразного») объяснения этого явления была выдвинута гипотеза о формировании над вершинами квазистационарных вихрей, которые п обеспечивают постоянный подъем вод, будучи настолько изолированными от окружающих вод, что внутри них образуются самостоятельные экологические системы. Действительно, подводный рельеф, усложняя в известной степени структуру набегающего потока, может способствовать образованию локальных вихрей, приводящих в конечном счете к обогащению поверхностных слоев биогенными элементами. Однако пространственно-временная устойчивость таких впхрей представляется довольно слабой, а их продолжительная изоляция — в высшей степени проблематичной. Ведь время, необходимое для развития фитопланктона, исчисляется несколькими сутками, а кормовая база рыб — мезопланктон и макропланктон — развивается намного дольше, соответственно около месяца и полугода.. Столь длительное существование над подводной возвышенностью неподвижных и существенно замкнутых вихревых образований почти невероятно: если влияние локального обогащения поверхностных вод биогенами и может отразиться на продуктивности зоопланктона и тем более па рыбопродуктивности, то произойдет это только в значительном удалении от места подъема вод. Таким образом, эффект увеличения биомассы кормовых организмов за счет локального «апвеллинга» может быть использован не бептопелагическими рыбами той подводной возвышенности, которая послужила причиной возникновения такого «шлейфового» пятна повышенной биомассы планктона, а только истинно пелагическими хищниками (например, тунцами и кальмарами). Поэтому гораздо более правомочным представляется другой подход, связывающий кормность поднятий с горизонтальным приносом (адвекцией) пищевых объектов набегающим потоком. Прямые исследования питания рыб, живущих над подводными возвышенностями, показывают, что их откорм происходит в основном за счет макропланктона, входящего в состав мигрирующих звукорассеивающих слоев (миктофиды, эвфаузииды и т. п.) и опускающегося днем на самую вершину горы (если, конечно, она лежит выше нормальной глубины дневного опускания этих слоев). Распространение. Географическое распространение океанических рыб, входящих в состав бентопелагического комплекса, еще не изучено в деталях, но представляется, что размеры их ареалов сильно варьируют. Некоторые из них весьма обширны — такие виды, как, например, эпимезобентальные антигония Antigonia capros и рыба-бекас Macrorhamphosus scolopax, мезобентальпые мягкоголовый долгохвост Malacocephalus laevis и низкотелый берикс Вегух splendens, мезобатибеитальные альдровандия Aldrovandia phalacra и антимора Antimora roslrata, батибентальные гладкоголов нарцет Narcetes sloimias и ксиелациба Xyelacyba myersi, батиабиссобентальные гладкоголов беллоция Bellocia koefoedi и батизавр Bathysaurus mollis, абиссобентальные слепой тифлонус Typhlonus nasus и эхиномакрурус Echinomacrurus mollis, обнаружены на соответствующих глубинах в тропических водах всех трех океанов. В то же время во всех вертикальных зонах есть и виды, которые известны из очень ограниченных участков и, по-видимому, на самом деле имеют очень узкое распространение (хотя представления об эндемизме могут основываться на недостаточных данных). Крайний случай почти «точечного» ареала демонстрирует распространение латимерии, обитающей только на склонах двух маленьках островов Коморского архипелага. Большинство других узколокальных эндемиков также обитает на вершинах подводных поднятий.
Известно, что дробность биогеографического деления бентали, достигающая наибольшей выраженности на шельфе и в верхней части склона (т. е. на глубинах залегания поверхностных и подповерхностных вод), уменьшается в нижней батиали и сводится к минимуму на ложе океана, где можно обнаружить только самые главные границы (те, которые разделяют регионы высшего ранга — области). Многими работами показано, что главные зоогеографические границы в мезобентали как бы продолжают вглубь границы, проходящие на шельфе. Действительно, анализ географического распространения рыб, населяющих склоны континентов и подводные поднятия, дает возможность выявить те же крупномасштабные регионы, что и в сублиторальной зоне. Так, Г. А. Го-ловань, рассматривая ареалы мезобентальных рыб, обитающих у западных побережий Африки и Eвропы, обнаружил в составе ихтиофауны 12 географических элементов и выделил четыре района — бореальный, северный субтропический, тропический и южный субтропический — в районировании шельфа.
Немногочисленность поимок многих видов батибентальных рыб сильно затрудняет анализ их географического распространения. Тем не менее представления о преобладании в этой зоне узколокальных ареалов, существовавшие до недавнего времени, не подкрепляются, как правило, новыми данными. Почти для всех видов, известных по многим нахождениям, характерно широкое распространение. Достаточно четкое разделение существует, однако, между фаунами тропических, умеренных и холодных вод. В тропической зоне выделяются экваториальный и широкотропический типы ареалов с циркумглобальным, индовестпацифическим, атлантическим и восточно-тихоокеанским подтипами. Умеренноводные ареалы подразделяются на потальные и бореальные, а холодноводные — на антарктические и арктические.
Что касается абиссобентальных и хадальных рыб, все они известны по немногим, часто единичным находкам, что сильно затрудняет рассмотрение их зоогеографии. Нужно отметить, впрочем, что на ложе океана некоторые виды уже сейчас найдены в далеко разобщенных районах (в двух или трех океанах встречены Bathymicrops regis, Ipnops meadi, Holcomycteronus profundissimus, Abyssobrotula galatheae и др.), и это позволяет предполагать их широкое распространение.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://aqualib.ru/ 'Подводные обитатели - гидробиология'